Введение
1.1.
Понятие
об анатомии
Анатомия животных –
раздел морфологии, изучающий развитие и строение организма животных в связи с функцией,
воздействием окружающей среды и наследственными факторами.
Задачи анатомии:
· Выявить
закономерности строения животного
· Использовать
эти закономерности в технологиях производства животноводства.
· Изучить
коррелятивное влияние изменяющегося органа на другие органы.
· Изучить
влияние внешних факторов на формообразование.
Значение
анатомии:
· Общеобразовательное
· Прикладное
Анатомия
– одна из самых древних биологических наук. Только из-за колоссальной сложности
строения организма и его функций, произошло разделение на анатомию (изучает
строение) и физиологию (изучает функцию).
Существует
несколько видов анатомии:
Макроскопическая –
наблюдение строения и сложные связи организма методом препарирования;
Микроскопическая –
наблюдение строения организма с помощью микроскопа;
Макро- микроскопическая –
пограничная зона между макро и микроскопической анатомией;
Описательная анатомия –
подробно характеризует строение каждого органа;
Системная – в
организме согласно главным функциям выделены системы и аппараты (системы:
костная, мышечная, общего кожного покрова, кровеносная, лимфатическая, нервная,
аппараты: пищеварительный, дыхательный, мочеполовой);
Видовая –
описывающая анатомию конкретных видов (человека, лошади, птицы и т.д.);
Топографическая –
изучает взаиморасположение органов (синтопия) и выносит проекцию органа на
скелет (скелетотопия);
Породная –
изучающая особенности строения органов у различных пород животных;
Возрастная –
строение различных органов в различные возрастные период;
Пластическая –
изучает внешние формы тела, пропорции, соотношение частей при различных
положениях;
Функциональная –
изучает строение органа в связи с его функцией;
Сравнительная –
изучает сходство и различие в строении органов животных;
Эволюционная –
изучает историю развития животных (рис. 1)
1.2. История
развития анатомии
Периоды истории
развития анатомии:
n Начальный
(500 гг. до н.э.).
n научный
(400 гг. до н.э. – 1500 гг. н.э.).
n период
возрождения (16 – 17 вв.).
n микроскопический
(2-ая половина 16 – 17 вв.)
n сравнительно-эмбриологический
(с 20 в.)
Папирус, написанным египетским
врачом Имхотепом (ХХХ век до н.э.), Папирус Эберса (около
2550 г. до н.э.) - были описаны некоторые органы, приведены данные об их
функциях
Гиппократ
(460-377 гг. до н.э.) собрал и систематизировал наблюдения о строении тела
человека. Заблуждения: артерии содержат воздух, мозг человека имеет
консистенцию слизи и вырабатывает семя, которое через спинной мозг поступает в
половые органы. Он не отличал нервы от сухожилий и вены от артерий. Гиппократа
считают отцом медицины, он большое значение придавал изучению анатомии, считая
ее первоосновой медицины (рис. 2).
Аристотель
(384-322 гг. до н.э.). Считал, что форма живых существ построена по высшим
идеальным образцам, впервые описал яичники, третье веко, сердечные кости у
жвачных, отметил отсутствие желчного пузыря у лошади, а также заложил основу научной
классификации животных и сравнительной анатомии (рис. 3).
Герофил (304
г до н.э.) (Александрийская школа врачей, им разрешалось препарирование и
вскрытие людей, живосечение осужденных). Описал оболочки мозга и венозные
пазухи, желудочки мозга и их сосудистые сплетения, зрительный нерв и глазное
яблоко, двенадцатиперстную кишку и лимфатические сосуды брыжейки. Опроверг
заблуждение Аристотеля, что сердце есть орган мышления, признав эту роль за
мозгом (рис. 4).
Клавдий Гален из
Пергама (139-201 гг). Римская школа. Впервые начал читать курс анатомии. Подробно
описал мышцы и кости, семь пар черепно-мозговых нервов, стенку артерий,
желудка, кишечного тракта и матки. Очень ценны сведения о строении головного
мозга. 13 столетий пользовались, зачастую ошибочными, данными Галена (рис. 5).
Авиценна или
Абу Али ибн Сина(980-1037 гг.). Этот период характеризуется вторжением в Европу
арабских народов. Авиценной систематизированы и дополнены знания по анатомии и
физиологии, заимствованные из трудов Аристотеля и Галена, но развитие анатомии
в этот период тормозит диктатура церкви (рис.
6).
Леонардо да Винчи
(1452-1519 гг.). Эпоха Возрождения, в которой заложен фундамент научной
анатомии. Леонардо положил начало пластической анатомии. Им выполнено более 800
очень точных рисунков костей скелета, мышц, внутренних органов,
сопровождающихся заметками и описаниями. Он изучал пропорции тела человека,
функциональную анатомию двигательного аппарата, особенности детского и
старческого организма (рис. 7).
Габриэль Фаллопий
(1523-1562) дал первое обстоятельное описание развития и строения ряда органов.
Его открытия изложены в книге «Анатомические наблюдения». Описал, в частности,
полукружные каналы, клиновидные пазухи, троичный, слуховой и языкоглоточный
нервы, канал лицевого нерва, а также маточные трубы, называемые в его честь
фаллопиевыми. Свои открытия Фаллопий описал в труде «Opera genuina omnia» (3
тома, Франкфурт, 1600, и Венеция, 1606) (рис.
8).
Бартоламео Евстахий
(1520-1574) являлся одним из
основоположников научной анатомии, в основу которой им были положены
сравнительно-анатомические исследования органов человека и человеческого
зародыша, а также патологоанатомические вскрытия животных. Имя Евстахия носят
открытая им «Евстахиева труба» (лат. tuba Eustachii) — соединительная труба
между барабанной полостью и носоглоточным пространством, и valvula Eustachii —
полулунный клапан нижней полой вены (vena cava inferior). Изучал и описывал
строение др. органов. Создал «Анатомические таблицы» (38 рисунков),
опубликованные в 1714. В своих воззрениях — последователь К. Галена и противник
А. Везалия (рис. 9).
Андреас Везалий
(1570-1657 гг.) был не только новатором в развитии анатомической техники, но и
реформатором в области анатомической номенклатуры, первым опроверг знания
Галена, излагая свои собственные. В 25 лет получил звание доктора медицины, а в
28 лет закончил и опубликовал классический труд в 7 книгах «О строении
человеческого тела». В них он описал в систематическом порядке скелет, связки,
мышцы, сосуды, нервы, внутренности, мозг и органы чувств (рис. 10).
Уильям Гарвей
(1578-1657 гг). Экспериментируя на животных, открыл большой круг кровообращения
и заложил научные основы физиологии. Ему принадлежит большой труд по
эмбриологии животных (рис. 11).
Испанский
естествоиспытатель М. Сервет (1521-1553), а через 6 лет Р. Коломбо (1516-1559) впервые высказали мысль о переходе крови из
правой половины сердца в левую через легочные сосуды, т.е. открыли малый круг
кровообращения. Ни Коломбо, ни Сервет, по-видимому, не знали об открытии араба
Ибн-ан-Нафиса из Дамаска (XIII в.) о легочном круге кровообращения (рис. 12).
Микроскопический
этап связан с Ренье де Грааф, Марчелло Мальпиги, Антони ван
Левенгук, Шумлянским, они описали клеточное строение животных, сделали
много открытий в строении капиллярных сосудов кожи, почек, трубчатых органов. С
именами этих ученых связаны названия некторых органов, структурных частей
органов организма человека и животных (рис.
13).
Основателем клеточной теории
строения организма явился немецкий ученый Шван Т. (1810-1882). Он
показал, что основной составной частью растительных и животных организмов
является клетка, установив тем самым общность строения органических форм
материи (рис. 14).
Прогресс русской науки
связан с многогранной деятельностью гениального ученого М.В. Ломоносова
(1711-1765 гг.), который уделял большое внимание развитию анатомии и физиологии
(рис. 15).
Н.И. Пирогов
(1810-1881). Участвовал в создании первого в России Анатомического института.
Им предложен метод изучения топографии внутренних органов на распилах
замороженных трупов. Изучал анатомию кровеносных сосудов, нервов, мышц и фасций
(рис. 16).
Эволюционный этап связан с:
Ч. Дарвиным
(1809-1882 гг.). Он обосновал изменчивость видов животных в процессе
приспособления их к условиям существования, единство животного мира и установил,
что человек возник в процессе эволюции от человекообразных обезьян. Благодаря
Дарвину возникла наука антропология (рис.
17).
В. О. Ковалевским
(1842-1882), он соединил эволюционно-морфологические данные с палеонтологией и
на основе этого написал «Палеонтологию лошадей» (рис. 18).
А. Н. Северцовым
(1866-1936), он разработал морфобиологическую теорию эволюционного процесса и
теорию филэмбриогеназа (рис. 19).
П.Ф Лесгафт
(1837-1909 гг.) является основоположником отечественной функциональной и
теоретической анатомии. Изучал закономерности строения сосудистой, мышечной и
костной систем организма, исследовал зависимость движений от формы суставных
поверхностей костей, взаимосвязь структуры и функции органов, одним из первых
применил экспериментальный метод на животных и методы математического анализа (рис. 20).
В.Н. Жеденов (1908-1962)
– основатель Оренбургской школы анатомов, совместно со своими учениками
разработали эволюционную морфологию сердца и легких млекопитающих (рис. 21).
Юрий Федорович Юдичев (1931-2000 гг.) — заслуженный деятель науки Российской Федерации,
академик МАН ВШ, доктор биологических наук, профессор, заведующий кафедрой
анатомии домашних животных Института ветеринарной медицины Омского
государственного аграрного университета, опубликовал более 160 научных работ, подготовил
десятки аспирантов, многие из которых в последствии стали кандидатами и
докторами наук, его имя включено в «Пять тысяч самых влиятельных людей мира» (рис. 22).
А.Ф. Климов (1878-1940 гг.) Заслуженный
деятель науки РСФСР, лауреат Государственной премии, доктор биологических наук
профессор - основатель московской школы и анатомического музея (рис. 23).
Продолжили работу московской
школы: профессор Ирина Владимировна Хрусталева (1919-2011), профессор Слесаренко
Наталья Анатольевна – заслуженный деятель науки РФ, почетный работник
высшего профессионального образования РФ, доктор биологических наук, зав.
кафедрой, декан ФВМ (рис. 24).
1.3. Объекты и
методы изучения анатомии
Объектами изучения анатомии
являются различные виды животных (рис 25).
Методы изучения анатомии:
I группа (рис. 26)
n рассечения,
или препарирования;
n вымачивания
трупов;
n распилов
замороженных трупов;
n наливки
или инъекции (заполнения) органов;
n коррозии
или разъедания;
n макро-
микроскопический;
n электронной
микроскопии.
II группа (рис. 27)
n рентгенологический и его модификации (рентгеноскопия,
рентгенография, рентгенокимография);
n соматоскопический;
n соматометрический,
или антропометрический;
n эндоскопии
внутренних органов;
n радиоизотопный;
n ультразвуковой
эхолокации с доплерографией;
n парамагнитного
резонанса.
1.4. Понятие об
органе, организме, системах и аппаратах
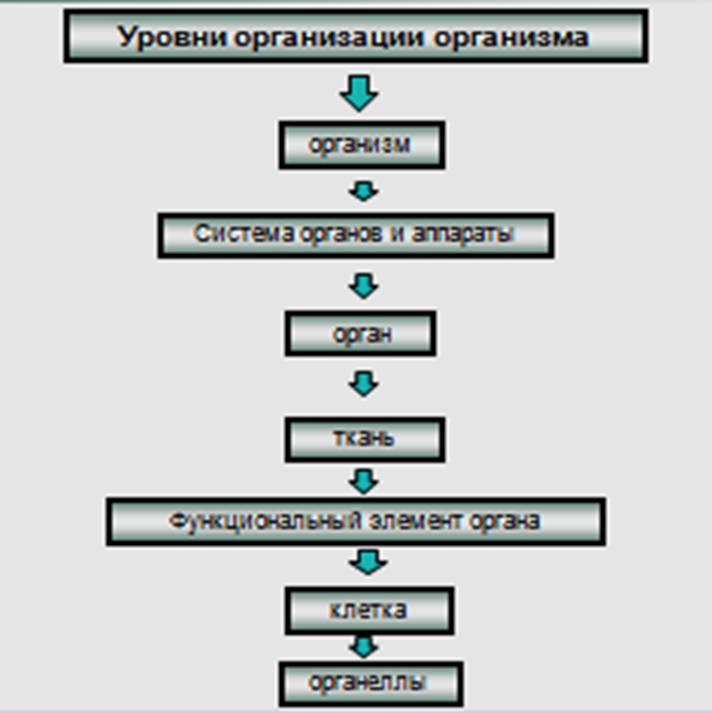
Организм - целостная
биотканевая динамичная система, имеющая иерархическое строение, элементы
которой функционируют как единое целое (рис. 28).
Система –
совокупность органов, имеющих общее происхождение, тесную морфологическую
взаимозависимость и выполняющих строго определенную функцию (рис. 29)
Аппарат -
совокупность органов, которые не смотря на различие в своем строении,
топографии и происхождении, объединены между собой для обеспечения какого-либо
определенного жизненно важного процесса в организме (рис.
30).
Орган – ориентированный в пространстве структурно-тканевой
комплекс, имеющий специфику строения и функционирования. Орган состоит из
нескольких тканей, одна из которых доминирует и определяет его функциональное значение (рис.
31).
Различают
следующие виды органов:
* паренхиматозные
(почки, печень) для них характерна строма и паренхима),
* трубкообразные
органы (пищевод, трахея),
* рудиментарные,
провизорные (переменные),
* атавистические,
дифинтивные (постоянные),
* аналогичные,
гомологичные, гомотипичные (органы, занимающие одно положение, и
симметрично похожие друг на друга),
* гомодинамные
(органы, построенные по определённому плану: позвонки, грудная и свободная
тазовая конечность.
Признаки органа:
1. форма.
2. Строение.
3. Происхождение (развитие).
4. Функции.
5. Положение в организме
(топография).
Органы объединены в три
группы:
1. Группа соматическая:
- скелет (кости и их
соединения)
- мускулатура (скелетная)
- кожный покров и его производные.
2. Группа
висцеральная (спланхническая):
- пищеварительный аппарат;
- дыхательный аппарат;
- мочеполовой аппарат:
органы выделения и размножения
3. Интегрирующая группа:
- сердечнососудистая система
- эндокринный аппарат
- нервная система
- органы чувств
Ткань –
совокупность более менее однородных клеток и их производных, объединённых
общностью строения, функций и происхождения.
Эпителиальная (рис. 32)
n Плоский
эпителий. Поверхность клеток гладкая. Клетки плотно примыкают друг
к другу (поверхность кожи, ротовая полость, пищевод, альвеолы, капсулы
нефронов)
Функция:
Покровная, защитная, выделительная (газообмен, выделение мочи)
n Железистый
эпителий. Железистые клетки вырабатывают секрет (железы кожи,
желудок, кишечник, железы внутренней секреции, слюнные железы)
Функция:
Выделительная (выделение пота, слез), секреторная (образование слюны,
желудочного и кишечного сока, гормонов)
n Мерцательный
(реснитчатый) эпителий. Состоит из клеток с многочисленными волосками(реснички)
(дыхательные пути)
Функция:
Защитная (реснички задерживают и удаляют частицы пыли)
Соединительная (рис. 33)
n Плотная
волокнистая Группы волокнистых, плотно лежащих клеток
без межклеточного вещества (собственно кожа, сухожилия, связки, оболочки
кровеносных сосудов, роговица глаза) Функция: Покровная, защитная,
двигательная
n Рыхлая
волокнистая Рыхло расположенные волокнистые клетки,
переплетающиеся между собой. Межклеточное вещество бесструктурное (подкожная
жировая клетчатка, околосердечная сумка, проводящие пути нервной системы)
Функция:
Соединяет кожу с мышцами, поддерживает органы в организме, заполняет промежутки
между органами. Осуществляет терморегуляцию тела
n Хрящевая
Живые
круглые или овальные клетки, лежащие в капсулах, межклеточное вещество плотное,
упругое, прозрачное (межпозвоночные диски, хрящи гортани, трахеи, ушная
раковина, поверхность суставов)
Функция:
Сглаживание трущихся поверхностей костей. Защита от деформации дыхательных
путей, ушных раковин
n Костная
Живые клетки с длинными отростками, соединенные между собой, межклеточное
вещество – неорганические соли и белок оссеин (кости скелета) Функция:
Опорная, двигательная, защитная
n Кровь
и лимфа Жидкая соединительная ткань, состоит из форменных
элементов (клеток) и плазмы (жидкость с растворенными в ней органическими и
минеральными веществами – сыворотка и белок фибриноген) (кровеносная система
всего организма)
Функция:
Разносит О2 и питательные вещества по всему организму. Собирает СО2 и продукты
диссимиляции. Обеспечивает постоянство внутренней среды, химический и газовый
состав организма. Защитная (иммунитет). Регуляторная (гуморальная)
Мышечная (рис. 34)
n Поперечно–полосатая
Многоядерные клетки цилиндрической формы до 10 см длины, исчерченные
поперечными полосами (скелетные мышцы, сердечная мышца)
Функция:
Произвольные движения тела и его частей, мимика лица, речь. Непроизвольные
сокращения (автоматия) сердечной мышцы для проталкивания крови через камеры
сердца. Имеет свойства возбудимости и сократимости
n Гладкая
Одноядерные
клетки до 0,5 мм длины с заостренными концами (стенки пищеварительного тракта,
кровеносных и лимфатических сосудов, мышцы кожи)
Функция:
Непроизвольные сокращения стенок внутренних полых органов. Поднятие волос на
коже
Нервная (рис. 35)
Нервные клетки (нейроны) Тела нервных клеток, разнообразные по форме и величине,
до 0,1 мм в диаметре Образуют серое вещество головного и спинного мозга Высшая
нервная деятельность. Связь организма с внешней средой. Центры условных и
безусловных рефлексов. Нервная ткань обладает свойствами возбудимости и
проводимости
Короткие отростки
нейронов – древовидноветвящиеся дендриты Соединяются
с отростками соседних клеток Передают возбуждение одного нейрона на другой,
устанавливая связь между всеми органами тела
Нервные волокна – аксоны (нейриты) – длинные выросты нейронов до 1,5 м длины. В
органах заканчиваются ветвистыми нервными окончаниями
Нервы периферической
нервной системы, которые иннервируют все органы тела Проводящие
пути нервной системы. Передают возбуждение от нервной клетки к периферии по
центробежным нейронам; от рецепторов (иннервируемых органов) – к нервной клетке
по центростремительным нейронам.
Вставочные нейроны передают возбуждение с центростремительных (чувствительных) нейронов на
центробежные(двигательные)
Клетка – наименьшая обособленная частичка организма с ядром
внутри.
Органеллы – постоянно
присутствующие, специализированные структурные элементы цитоплазмы. Благодаря
им поддерживается важнейшая функциональная и структурная специализация клетки (рис. 36)
1.5. Основные
проявления жизни
·
Реактивность
(раздражимость) – свойство живой материи
не только воспринимать раздражения, исходящие из внешней или внутренней среды,
но и отвечать на эти раздражения соответствующей реакцией.
Осуществляется нервной системой, эндокринной и органами чувств.
·
Обмен веществ и энергии. Осуществляется за счёт системы пищеварения и дыхания при активном участии
сердечнососудистой системы, которая транспортирует от органов пищеварения и
дыхания ко всем органам и тканям питательные вещества и кислород, а забирает от
них продукты метаболизма к органам выделения. Теплорегуляция осуществляется
мышечной системой, работой различных органов и кожей. Регуляция и адаптивная
перестройка интенсивности обменных процессов происходят под контролем нервной
системы через посредство желез внутренней секреции и сердечнососудистой
системы.
·
Размножение обеспечивает
преемственность жизни на нашей планете и сохранение определенной численности
видов. При ослаблении этой способности или с ее прекращением виды обречены на
вымирание. Для позвоночных свойственно половое размножение. Половые клетки,
вырабатываемые в половых железах, содержат в себе всю необходимую
генетическую информацию, передаваемую
будущему потомству при оплодотворении. Осуществляется, за счет половой
системы. Процесс размножения находится под воздействием нервной и гуморальной
регуляции
1.6. Общие
принципы строения организма млекопитающих
· Одноосность
(биполярность) –
это наличие двух полюсов тела: головного (краниального) и хвостового
(каудального).
· Трехслойное строение
зародыша –
зародыш состоит из трех слоев: энтодермы, эктодермы, мезодермы.
· Антимерия
(двусторонняя, билатеральная симметрия) – выражается в сходстве по строению правой и
левой половин тела, поэтому большинство органов парные (глаза, уши, легкие,
почки, грудные и тазовые конечности).
· Сегментарность
(метамерия)
– близлежащие участки тела (сегменты) близки по строению. У млекопитающих
сегментарность четко выражена в осевом скелете.
· Закон трубкообразного
строения. Все
системы и аппараты организма развиваются из трубок в виде трубок (нервная
система, пищеварительный аппарат)
· Расположение непарных
органов вдоль основной оси тела животного располагаютсянепарные органы
(сердце, печень, поджелудочная железа, трахея.
1.7. Плоскости и
направления в анатомии
В
теле животного проводят 3 плоскости: саггитальную, сегментальную и фронтальную (рис. 37).
n Саггитальная
плоскость (серединная) – lamina saggitalis (medialis) -это плоскость, проведённая вертикально вдоль
тела животного, разделив тело на 2 половины правую и левую. С этой плоскостью
связаны 2 направления : латеральное (lateralis)и медиальное (medialis) направления.
n Сегментальная
плоскость – lamina
segmentalis – это плоскость, проведённая вертикально поперёк тела, и
делящая его на переднюю и заднюю части. С этой плоскостью связаны 2
направления: краниальное ( "cranium" – голова) и каудальное (
"caudae" – хвост). В области головы с этой плоскостью связаны
направления: назальное – nasalis – к
носу, оральное (ростральное у свиней) – oralis (rostralis) – ко рту, аборальное – aboralis – назад.
n Фронтальная
плоскость – lamina
frontalis – это плоскость, проведённая горизонтально поперёк тела
животного, и делящая его на верхнюю и нижнюю части. Связаны направления:
дорсальное ( "dorsum" – спина) и вентральное ( "Venter" –
живот). В области конечностей: проксимальное – proximalis – вверх, дистальное – distalis – вниз; на кисти:
дорсальное – dorsalis –
вперёд, пальмарное, волярное – palmaris, volaris – назад; на стопе:
плантарное plantaris –
назад, дорсальное – вперёд.
1.8. Основные
законы биологического развития
1.8.1. Закон
исторического развития. Филогенез
Филогенез –
историческое развитие вида от одноклеточных до ныне существующих форм.
Принципы филогенеза по И.И.
Шмальгаузену:
· В процессе развития организма постоянно идет дифференцировка клеток и
тканей с одновременной их интеграцией. Дифференциация – это разделение между
клетками функций, одни участвуют в переваривании пищи, другие, как например,
эритроциты, в переносе кислорода. Интеграция – это процесс укрепления между
клетками, тканями взаимосвязей, которые обеспечивают организму целостность (рис. 38).
· Каждый орган имеет несколько функций, но одна из них является главной.
Остальные функции являются как бы второстепенными, запасными, но благодаря им,
орган имеет возможность преобразовываться. Например поджелудочная железа
выполняет две функции: эксреторную и инкреторную, а относится к пищеварительным
железам (рис. 39).
· При изменении условий жизни может произойти смена главной функции на
второстепенную и наоборот. Например печень в эмбриональном периоде выполняет
кроветворную функцию, а после рождения основная
функция – выработка желчи (рис. 40).
· В организме всегда наблюдаются два противоположных процесса:
прогрессивное развитие и регрессивное (редукция). Органы, которые утрачивают
свои функции, как правило, подвергаются редукции, т.е. постепенному
исчезновению. Иногда они сохраняются в виде рудимента (при сохранении
второстепенной функции) (рис. 41).
· Все изменения в организме
происходят коррелятивно, т.е. изменения в одних органах непременно ведут к
изменениям в других органах (рис. 42).
1.8.2. Закон
единства организма и среды
Организм без внешней среды,
поддерживающей его существование, невозможен (рис. 43).
Биологический прогресс у
животных в окружающей среде по А.Н. Северцову достигается
4 путями:
а)
путем ароморфоза, т.е. морфофизиологического прогресса, в результате которого
усложняется организация животного, и происходит общий подъем энергии
жизнедеятельности;
б)
путем идиоадаптации, т.е. частных полезных приспособлений, но при этом сама
организация животного не усложняется (губки, кишечнополостные);
в)
путем ценогенеза, т.е. эмбриональных приспособлений, которые развиваются только
у зародышей, а у взрослых исчезают (лягушка);
г)
путем общей дегенерации, т.е. упрощением организации и снижением интенсивности
жизнедеятельности, при этом интенсивно развивается половая система и пассивные
органы защиты, вследствие чего так же достигается победа в борьбе за существование
(паразитические плоские и круглые черви).
1.8.3. Закон
целостности и неделимости организма
Этот закон выражается в том,
что каждый организм является единым целым, в котором все органы и ткани
находятся в тесной взаимосвязи (рис. 44).
1.8.4. Закон
единства формы и функции
Форма и функция органа
образуют единое целое (рис. 45).
1.8.5. Закон
наследственности и изменчивости
В ходе возникновения и развития
жизни на Земле наследственность играла важную роль, обеспечивая закрепление
достигнутых эволюционных преобразований в генотипе. Она неразрывно связана с
изменчивостью. Благодаря наследственности и изменчивости стало возможным
существование разнообразных групп животных, многие из которых еще не изучены (рис. 46).
1.8.6. Закон
гомологичных рядов
гласит о том, чем ближе
генетические виды, тем больше они имеют сходных морфологических и
физиологических признаков (рис. 47).
1.8.7. Закон экономии материала и места
Согласно этому закону каждый
орган и каждая система построены так, чтобы при минимальной затрате
строительного материала они могли бы выполнять максимальную работу (рис. 48).
1.8.8. Основной
биогенетический закон (Бэра-Геккеля)
состоит в том, что
индивидуальное развитие организма (онтогенез) является коротким и быстрым
повторением (рекапитуляцией) важнейших этапов эволюции вида (филогенеза).
Например, закладка жаберных щелей у наземных позвоночных (рис.
49).
1.8.9.
Онтогенез, периоды, этапы и критические фазы развития организма животных в
онтогенезе
Онтогенез
("онто" – один, "генез" – развитие) – это жизненный период
животного от зиготы (слияние 2х гамет и двух половых клеток яйцеклетки и
спермотазоида) до смерти. Различают 2 периода:
внутриутрбный (пренатальный) и внеутробный (постнатальный) (рис. 50).
Законы индивидуального развития
по Л.П. Тельцову (2008)
n 1
закон: индивидуальное развитие состоит из 3 периодов –
эмбрионального, постнатального, зрелости. Каждый период включает несколько
этапов, стадий и критических фаз (КФ);
n II закон:
наследственность человека и животных реализуется по этапам развития. На каждом
этапе химический состав клеток, морфологические и физиологические возможности
тканей, органов и систем организма иные;
n III закон:
взаимодействие последовательных этапов протекает по принципу акселерации или ретардации.
Компенсация роста и развития прямо пропорциональна интенсивности воздействия в
следующем этапе и обратно пропорциональна возрасту;
n IV закон: КФ
развития организма выявляются на стыке этапов и характеризуются активацией
генов под влиянием внешних и внутренних факторов;
n V закон:
продолжительность критических фаз организма зависит от глубины перестройки в
последующем этапе;
n VI закон: на
каждом этапе развития имеются присущие только ему биологические ритмы;
n VII закон:
непрерывность развития организма обусловлена асинхронностью и гетерехронностью
составляющих его систем;
n VIII закон:
смена этапов онтогенеза обусловлена плавностью
n дефинитивных
тканей, органов и систем. На каждом этапе функционирует новая генерация тканей,
органов и систем.
В
критические фазы развития происходят:
- смена одного этапа развития организма
другим;
-
установка генетической программы на следующий этап;
-
подведение итогов прошедшему этапу;
-
десинхронизация биологических ритмов органов и систем организма;
-
повышение чувствительности тканей, органов и систем организма к лекарственным веществам и к
воздействию факторов внешней среды;
-
генетические мутации в клеточных дифферонах;
-
смена функций генераций временных и дефинитивных органов организма в
онтогенезе.
2.
Аппарат
движения (apparatus lokomotorus)
Одна из функций организма животных – изменение
положения частей тела, передвижение в пространстве. Движения происходят при
участии костей, выполняющих функции рычагов, и скелетных мышц, которые вместе с
костями и их соединениями составляют пассивную часть опорно-двигательного
аппарата, а мышцы, выполняющие функции сокращения и изменения положения костей,
- активную часть.
Прогрессивному
развитию млекопитающих способствовала их высокая двигательная активность, выразившаяся
в возникновении совершенных и очень разнообразных способов передвижения
(ходьба, бег, лазанье, плавание, полет) и адаптации к различным условиям
существования (наземное, наземно-древесное, воздушное, водное) (рис. 51).
2.1.
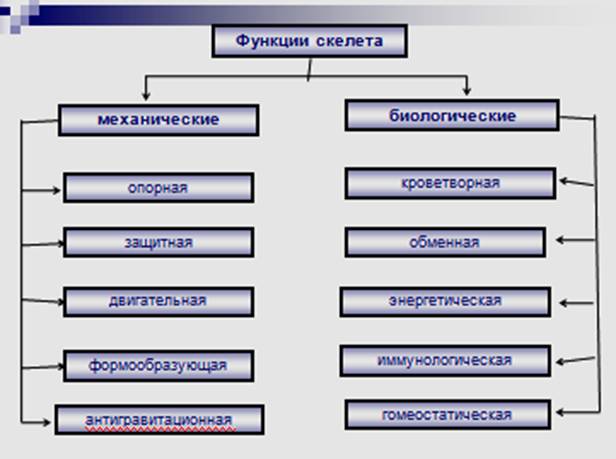
Характеристика скелета, принципы его строения
и деления на отделы. Роль скелета в
жизнедеятельности организма
Остеология –
("ост" – кость, "логос" – наука), раздел анатомии,
изучающий кости. В совокупности все кости формируют скелет
("skeleton" – высохший) – это совокупность костей, расположенных в
естественном порядке.
Характеризуя
скелет млекопитающих, отметим, во-первых, постоянное число шейных позвонков: у
подавляющего большинства видов этого класса, в том числе у китов и у жирафы,
шейный отдел содержит семь позвонков; только у ламантина (из отряда
американских неполнозубых) и ленивца число их колеблется между 6 и 10.
Другую
характерную особенность скелета млекопитающих составляет недоразвитие в их
плечевом поясе коракоидов, или вороньих костей, остатки которых прирастают к
лопаткам в виде коракоидных отростков; только у яйцекладущих млекопитающих -
утконоса и ехидны - в числе их древних, рептильных признаков коракоиды
сохраняются полностью. Таким образом, у высших млекопитающих связующим звеном между
лопатками и грудиной остается только пара ключиц, да и они отсутствуют у тех
животных, у которых конечности служат только для шага и бега и не приспособлены
для лазания, хватания или раскапывания земли (у копытных).
Череп.
Существенными особенностями характеризуется череп млекопитающих. Во-первых, он
сочленяется с позвоночником посредством двух затылочных мыщелков, что отличает
его от черепов рептилий и птиц и сближает с черепом амфибий.
У
млекопитающих имеется только одна височная (скуловая) дуга, которая по своей
природе не соответствует ни верхней, ни нижней скуловой дуге на черепе гаттерии
и крокодилов, - она образована скуловой костью и отростками челюстной и
чешуйчатой костей, тогда как у названных рептилий сочетание костей иное (у
современных ящериц сохранились только верхние дуги, у птиц - только нижние, а у
змей отсутствуют и те и другие). Любопытно отметить, что у черепах, в отличие
от других рептилий, имеется единственная скуловая дуга, образуемая теми же
костями, что и у млекопитающих.
Скелеты
крупных и мелких зверей. Как известно, скелет дает опору телу
животного и служит его механическим каркасом; поэтому скелет определяет общую
форму тела, которая в свою очередь связана с размерами животного и со способом
его передвижения (летучие мыши и киты).
У
крупных животных соотношение между мышечной силой и массой тела оказывается
чрезвычайно неблагоприятным, а это ведет к тому, что с увеличением размеров
должна меняться и форма мышц, которые становятся более утолщенными. Но раз
мышцы делаются более мощными и массивными, то для прикрепления их к
соответствующим костям скелета требуются и более широкие поверхности в виде
различных гребней и выростов на костях конечностей.
И
только у китов, которые целиком приспособились к водному образу жизни и которым
- в соответствии с законом Архимеда - приходится применять мышечную силу только
для передвижения, а не расходовать ее на поддержание тяжести своего тела,
увеличение размеров далеко оставило за собой и слонов и индрикотериев: для того
чтобы уравновесить на весах тушу одного синего кита, потребовалось бы поставить
на другую чашку весов 25 слонов.
Скелет
и моторика млекопитающих.
Присматриваясь
к тому, как перебегают мелкие зверьки, мы заметим, что и на коротких ножках они
передвигаются очень быстро. Достигается это тем, что в их передвижении
принимает активное участие и туловище: оно то сгибается, то с силой
распрямляется, и тогда зверек, оттолкнувшись задними ногами, выбрасывает тело
вперед и таким образом передвигается скачками.
Размеры таких скачков зависят не только от мускулатуры
задних конечностей (как, например, у лягушки), но и от мускулатуры самого
туловища: сгибаясь и распрямляясь, оно действует здесь наподобие стальной
пружины.
Если такую пружину мы сперва согнем, сжав ее концы между
пальцами, а затем отпустим и дадим ей распрямиться, она с силой отлетает от нас
прочь.
Такому способу передвижения соответствуют и особенности
скелета мелких млекопитающих (хотя бы того же лемминга). Их позвоночник
представляет собой легкий и гибкий стержень, на котором лишь слегка выступают
небольшие остистые отростки. Даже и при спокойной позе животного его
позвоночник находится в полусогнутом состоянии, как бы уже наготове для
активного сгибания и разгибания при передвижении зверька; это дает ему
возможность и при малом росте преодолевать бесчисленные препятствия, стоящие на
его пути, - кочки и рытвины, поваленные стволы деревьев, мелкие кустарнички.
Цепкие лапки с когтями на пальцах позволяют многим таким зверькам взбираться по
стволам вверх и вести древесный образ жизни (белка, куница).
Животные пальцеходящие и копытные. С увеличением
размеров четвероногого животного его костяк становится более массивным и
позвоночник с выступающими на нем остистыми отростками мало-помалу утрачивает
подвижность (сравните в этом отношении кошку и собаку). Быстрота передвижения
достигается здесь удлинением конечностей. Животные становятся не только
абсолютно, но и относительно более высокими на ногах; при этом на ногах
уменьшается поверхность соприкосновения их с землей. Животное при ходьбе и беге
опирается не на всю стопу, а только на пальцы,- как большинство более или менее
крупных хищников (животные пальцеходящие), или же только на концы пальцев,
"обутые" в этих случаях твердыми роговыми копытами (животные копытные).
О том, что сокращение опорной поверхности конечностей
способствует увеличению быстроты бега, свидетельствует наш житейский опыт:
пускаясь бежать, мы, существа стопоходящие, становимся на пальцы (на цыпочки),
а символом медлительности у нас недаром служит брюхоногий моллюск улитка,
ползущая на огромной "подошве".
Сокращение опорной поверхности достигается перенесением
ее со всей стопы на нижнюю сторону пальцев, а затем на копыта, одевающие
последние фаланги пальцев. Простейшее наблюдение на собственной руке покажет,
как в ходе исторического процесса уменьшения поверхности опоры становятся
излишними боковые пальцы, начиная с большого.
Важно отметить, что одностороннее приспособление
конечностей только к шагу и бегу ведет за собой и утрату ключиц, теряющих в
этом случае свое механическое значение. Чтобы это положение стало более ясным,
можно провести такое самонаблюдение. Обопритесь обеими руками на сиденье стула.
В таком положении руки будут служить подобием передних конечностей
четвероногих. Вы почувствуете, что ваши плечевые кости, упираясь в лопатки,
давят на них снизу вверх, причем на долю ключиц, идущих к грудине, не
приходится никакой нагрузки (иное дело, если мы будем разводить руки в стороны,
поднимать стул и т. д.).
Значение стопоходности и пятипалости. Однако из того, что
сказано здесь о пальцеходящих и копытных четвероногих, было бы неправильным
вывести заключение, что стопоходность свойственна только примитивным группам
млекопитающих, и притом только зверькам мелких размеров. Если быстрый бег не
служит животному основным способом его передвижения, если животному не
приходится постоянно применять его либо в погоне за добычей, либо для спасения
своей жизни и особенно если животное обитает на деревьях, то и стопоходность и
пятипалость (по крайней мере на задних конечностях) оказываются для него
полезными или даже необходимыми в связи с условиями его жизни.
Классический пример стопоходного животного - косолапый
мишка при кажущейся его неуклюжести способен к очень разнообразным движениям.
Как раз именно стопоходность и дает медведю возможность легко подниматься на
задние ноги и передвигаться в таком положении, не теряя равновесия; она же, при
наличии пяти когтистых пальцев, позволяет ему взбираться на деревья. Благодаря
этим природным данным в руках опытных дрессировщиков медведи становятся
искусными цирковыми артистами и выступают на арене в качестве велосипедистов,
эквилибристов, борцов и боксеров.
Только на основе стопоходности и пятипалости могли
развиться крылья летучих мышей и цепкие хватательные конечности прирожденных
древолазов-обезьян, и, наконец, только прочно встав на всю ступню "задних
ног" и освободив от работы по передвижению пятипалые руки, наши
обезьяноподобные предки смогли очеловечиться, начав изготовлять и применять
различные орудия.
Неожиданное
открытие сделали итальянские ученые, когда им удалось из древесины с помощью
химических реакций создать кость, обладающую способностью срастаться с
собственными костями млекопитающих и выполнять все необходимые функции (рис. 52).
Как
сообщила сегодня британская радиостанция Би-би-си, сделанные из ротанговой
пальмы кости были пересажены в конечности овцам и сейчас практически ничем не
отличаются от естественных. Секрет превращения древесины в костную ткань заключается
в ее специальной обработке, отмечают ученые из лаборатории Айтек под
итальянским городом Болонья.
Первоначально
из ротанга вырезают необходимую кость или ее фрагмент. Затем древесину
нагревают в специальном растворе, содержащем уголь и кальций. После этого
деревянные кости помещают в специальную камеру с фосфатным раствором и держат
под сильным давлением. На эти операции требуется всего 10 дней.
Обработанную
таким образом древесину пересаживают животным. Би-би-си сообщает, что, как
показали опыты, костный ротанг получает способность абсорбировать в живом
организме биологически активные элементы и срастается с костью, не оставляя
даже нароста.
Как
заявила руководитель работ профессор Анна Тампьери, фактически ротанг
превращается в настоящую костную ткань.
Она
также сообщила, что в ближайшие 5 лет будут созданы ротанговые кости и для
человека.
Весь скелет
подразделяется на осевой и перифирический. Осевой скелет: скелет головы, скелет
шеи (позвоночник, рёбра, грудная кость), скелет туловища, крестца и хвоста.
Перифирический:
склет грудной и тазовой конечностей. Скелет грудной и тазовой конечностей
делится на пояс свободной конечности и свободную конечность.
Скелет животных.
Остеология –
раздел анатомии, изучающий кости. ("ост" – кость, "логос" –
наука). В совокупности все кости формируют скелет ("skeleton" –
высохший) – это совокупность костей, расположенных в естественном порядке (рис. 53).
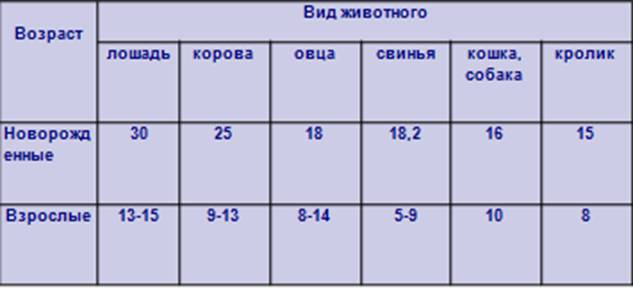
Масса скелета у домашних
животных в % к массе тела (По Жеденову В.Н.)
2.2.
Фило- и онтогенез скелета животных
Филогенез и онтогенез (мезенхимное происхождение) скелета протекает
в 3 этапа (стадии):
1. перепончатый (соединительнотканный);
2. хрящевой
3. костный
Беспозвоночные. Среди
простейших радиолярии обладают сложным, геометрически правильным кремневым
скелетом, а фораминиферы защищены известковыми раковинами своеобразной формы.
Скелеты губок могут быть построены из трех различных материалов: извести,
рогоподобного белка спонгина и кремнезема. Известь и спонгин иногда
комбинируются, но у стеклянных губок скелет чисто кремневый. У кишечнополостных
скелет встречается редко, если не считать кораллов, у которых он образован как
наружными, так и внутренними известковыми структурами. Известняки коралловых
рифов - это в основном отложения скелетов отмерших кораллов. У всех примитивных
групп скелет выполняет поддерживающую и защитную роль, но не используется для
локомоции. Плоские и круглые черви его лишены. Некоторые кольчатые черви живут
в известковых трубках, образованных их собственными выделениями. У червей
разных типов встречаются щетинки, которые считаются скелетными структурами.
Известковые раковины моллюсков - это в основном наружные образования;
исключение составляет внутренняя раковина каракатицы. Слизни и осьминоги
скелета лишены. Для членистоногих характерен составной скелет, покрывающий
снаружи все их тело, включая антенны (усики) и ноги. Он состоит из углевода
хитина, а у ракообразных может содержать большие количества кальция. Хитиновый
панцирь, развивающийся в эмбриогенезе из эктодермы, представляет собой мертвое
образование и расти не может, поэтому, увеличиваясь в размерах, все
членистоногие периодически сбрасывают наружный слой скелета (линяют). Круглые
черви по мере роста также неоднократно меняют свой жесткий наружный покров,
называемый кутикулой (рис. 54).
Позвоночные. Скелет позвоночных
образован не только костями: он включает в себя хрящ и соединительную ткань, а
иногда в его состав входят различные кожные образования. У позвоночных принято
выделять осевой скелет (череп, хорда, позвоночник, ребра) и скелет конечностей,
включающий их пояса (плечевой и тазовый) и свободные отделы. У ланцетников есть
хорда, но ни позвонков, ни конечностей нет. Змеи, безногие ящерицы и червяги
лишены скелета конечностей, хотя у некоторых видов первых двух групп
сохраняются их рудименты. У угрей соответствующие задним конечностям брюшные
плавники исчезли. У китов и сирен никаких внешних признаков задних ног также не
осталось.
Череп. По происхождению различают три
категории костей черепа: замещающие хрящ, покровные (накладные, или кожные) и
висцеральные. Беспозвоночные лишены структуры, сравнимой с черепом позвоночных.
У полухордовых, оболочников и головохордовых никаких признаков черепа нет. У
круглоротых череп хрящевой. У акул и их родичей в нем, возможно, когда-то
присутствовали кости, но сейчас его коробка представляет собой единый монолит
из хряща без швов между элементами. У костных рыб в черепе больше различных
костей, чем у представителей любого другого класса позвоночных. У них, как и
всех вышестоящих групп, центральные кости головы закладываются в хряще и
замещают его, а следовательно, гомологичны хрящевому черепу акул. Покровные
кости возникают как известковые отложения в дермальном слое кожи. У некоторых
древних рыб они представляли собой пластины панциря, защищающего мозг,
черепно-мозговые нервы и находящиеся на голове органы чувств. У всех высших
форм эти пластины мигрировали в глубину, включились в исходный хрящевой череп и
образовали новые кости, тесно связанные с замещающими. Почти все наружные кости
черепа происходят из дермального слоя кожи. Висцеральные элементы черепа -
производные хрящевых жаберных дуг, возникших в стенках глотки при развитии у
позвоночных жабр. У рыб две первые дуги видоизменились и превратились в
челюстной и подъязычный аппарат. В типичных случаях у них сохраняются еще 5
жаберных дуг, но у некоторых родов их число сократилось. У примитивной
современной акулы семижаберника (Heptanchus) позади челюстной и подъязычной дуг
жаберных дуг целых семь. У костных рыб челюстные хрящи облицованы
многочисленными покровными костями; последние образуют также жаберные крышки,
защищающие нежные жаберные лепестки. В ходе эволюции позвоночных исходные хрящи
челюстей неуклонно редуцировались, пока не исчезли полностью. Если у крокодилов
остаток первоначального хряща в нижней челюсти облицовывают 5 парных покровных
костей, то у млекопитающих из них остается лишь одна - зубная, которая
полностью формирует скелет нижней челюсти. Череп древних амфибий содержал
тяжелые покровные пластинки и был похож в этом отношении на типичный череп
кистеперых рыб. У современных земноводных как накладные, так и замещающие кости
сильно редуцированы. В черепе лягушек и саламандр их меньше, чем у других
позвоночных с костным скелетом, причем в последней группе многие элементы
остаются хрящевыми. У черепах и крокодилов кости черепа многочисленны и плотно
сращены между собой. У ящериц и змей они относительно невелики, причем наружные
элементы разделены широкими промежутками, как у лягушек или жаб. У змей правая
и левая ветви нижней челюсти весьма свободно соединены между собой и с черепной
коробкой эластичными связками, что позволяет этим рептилиям заглатывать
относительно крупную добычу. У птиц кости черепа тонкие, но очень твердые; у
взрослых особей они срослись настолько полно, что несколько швов исчезли. Очень
велики глазничные впадины; крыша относительно огромной мозговой коробки
образована тонкими покровными костями; легкие челюсти покрыты роговыми чехлами.
У млекопитающих череп тяжелый и включает мощные челюсти с зубами. Остатки
хрящевых челюстей переместились в среднее ухо и образовали его косточки -
молоточек и наковальню.
Впервые скелетные
образования возникли у простейших. Скелет радиолярий (лучевиков), состоящий у
большинства этих созданий из кремнезёма, придаёт им необычайную красоту. Можно
выделить два направления дальнейшей эволюции скелета. Первое — формирование наружного
скелета (например, раковины моллюсков, панцирь ракообразных, твёрдый
покров насекомых). Второе — развитие внутреннего скелета (у радиолярий,
губок, позвоночных животных).
Долгое время на суше
господствовали исключительно животные с наружным скелетом. Он весьма удачно
защищает его обладателей от нападения врагов, служит опорой их телу, помогает
передвижению. Иногда ему достаётся самая неожиданная роль. Корабельному червю
(один из видов моллюсков) остаток раковины помогает просверливать ходы в
древесине, которой он питается.
Основной материал для
раковин моллюсков и фораминифер, скелета полипов — известь, т. е. карбонат кальция.
(Из почти чистого карбоната кальция состоят мел и скорлупа куриных яиц.) Но
вещество это довольно плотное, тяжёлое. Животные с внешним известковым
скелетом на суше весьма неповоротливы. Медлительность улиток даже вошла в
поговорку.
Самый совершенный
наружный скелет — у членистоногих. Они сумели «найти» для него более удачный и
лёгкий материал — хитин Для большей прочности хитин может пропитываться той же
известью. Твёрдые щитки, покрывающие тело животного, соединяются друг с другом
более мягкими перепонками. Это позволило членистоногим иметь характерные для
них членистые конечности «на шарнирах», столь незаменимые при передвижении по
суше. Их скелеты напоминают суставчатые рыцарские латы. Мышцы крепятся к
такому скелету изнутри (что можно увидеть при разделывании клешней рака). В
конечном итоге благодаря всем этим приспособлениям членистоногие сумели
завоевать сушу.
К сожалению, наружный
скелет членистоногих имеет и ряд недостатков. Он не может нарастать по мере
роста животного, поэтому мешает росту. Чтобы расти, животному приходится время
от времени покидать свой скелет — претерпевать линьку. Наблюдательные
читатели нередко могли заметить висящую на паутине пустую оболочку паучьих лап
— паук покинул свой скелет. Сразу после линьки животное быстро растёт, пока не
затвердеют его новые покровы. Но в этот период оно легко уязвимо для хищников.
Лишённое опоры, оно вдобавок почти не может двигаться (в особенности если живёт
на суше). На время линьки животные обычно прячутся в укромные места.
И ещё одно
несовершенство наружного скелета. С уверенностью можно сказать, что муравьи
размером с быка и тому подобные гигантские насекомые могут существовать лишь в
произведениях писателей-фантастов. У гигантской стрекозы, жившей в
каменноугольном периоде палеозойской эры, размах крыльев достигал
Наружный скелет. У позвоночных всех классов так или иначе представлен наружный
скелет. Головные пластины у щитковых (вымерших бесчелюстных), древних рыб и
амфибий, так же как чешуи, перья и волосы высших четвероногих, представляют
собой кожные образования. Таков же по происхождению панцирь черепах - высоко
специализированное скелетное образование. Их кожные костные пластинки
(остеодермы) приблизились к позвонкам и ребрам и слились с ними. Примечательно
то, что плечевой и тазовый пояса параллельно этому сместились внутрь грудной
клетки. В гребне на спине у крокодилов и панцире броненосцев находятся костные
пластинки того же происхождения, что и панцирь черепах.
Более перспективным в
эволюционном плане оказался путь развития не наружного, а внутреннего скелета.
Зарывшись в песок на
дне моря, процеживая из воды съедобные частицы, проводит свою жизнь небольшое
(5—
В ходе эволюции скелет
позвоночных постепенно утрачивает излишнюю гибкость и становится всё более
твёрдым. У хрящевых рыб (акул и скатов) скелет состоит ещё только из хрящевой
ткани. Хрящ лучше приспособлен для роста, чем твёрдая костная ткань, —
он может нарастать, не меняя пропорций. Зато костная ткань более прочна и
тверда. Позвоночник, или
позвоночный столб, есть у всех хордовых, за исключением бесчерепных и
оболочников. В эмбриональном развитии ему всегда предшествует хорда, которая
сохраняется пожизненно у ланцетника и круглоротых. У рыб она окружается
позвонками (у акул и их ближайших родственников - хрящевыми) и выглядит
четковидной. У млекопитающих сохраняются лишь рудименты хорды в межпозвоночных
дисках. Хорда не преобразуется в позвонки, а заменяется ими. Они возникают в
ходе эмбрионального развития как изогнутые пластинки, постепенно окружающие
хорду кольцами и по мере роста почти полностью ее вытесняющие.
У птиц и рептилий череп причленяется
к позвоночнику при помощи одного своего мыщелка (суставного бугорка). У
современных амфибий и всех млекопитающих для этого используются два мыщелка,
расположенные по бокам от спинного мозга (рис. 55).
Скелет конечностей. Конечности
четвероногих развились из парных плавников кистеперых рыб, в скелете которых
были элементы, гомологичные костям плечевого и тазового пояса, а также передних
и задних ног. Первоначально в плечевом поясе было по крайней мере пять
отдельных окостенений, однако у современных животных их обычно всего три:
лопатка, ключица и коракоид. Почти у всех млекопитающих коракоид редуцирован,
прирос к лопатке или вообще отсутствует. У некоторых зверей лопатка остается
единственным функциональным элементом плечевого пояса. Тазовый пояс включает
три кости: подвздошную, седалищную и лобковую. У птиц и млекопитающих они
полностью слились между собой, в последнем случае образовав т.н. безымянную
кость. У рыб, змей, китов и сирен тазовый пояс не прикреплен к позвоночнику, в
котором поэтому нет типичных крестцовых позвонков. У некоторых животных как
плечевой, так и тазовый пояс включают в себя добавочные кости. Кости передней
свободной конечности у четвероногих в принципе такие же, что и в задней, но
называются по-другому. В передней конечности, если считать от туловища, сначала
идет плечевая кость, за ней лучевая и локтевая кости, затем запястные, пястные
и фаланги пальцев. В задней конечности им соответствуют бедренная, затем
большая и малая берцовые, предплюсневые, плюсневые кости и фаланги пальцев.
Исходное число пальцев - по 5 на каждой конечности. У амфибий на передних лапах
всего по 4 пальца. У птиц передние конечности преобразованы в крылья; кости
запястья, пясти и пальцев сокращены в количестве и частично сращены между
собой, на ногах утрачен пятый палец. У лошадей остался только средний палец.
Коровы и их ближайшие родственники опираются на третий и четвертый пальцы, а
остальные утрачены или редуцированы. Копытные животные передвигаются на
кончиках пальцев и называются фалангоходящими. Кошки и многие другие звери при
ходьбе опираются на всю поверхность пальцев и относятся к пальцеходящему типу.
Медведи и человек при движении прижимают к грунту всю подошву и называются
стопоходящими (рис. 56).
В процессе онтогенеза скелет проходит три стадии
своего развития и формирования. На ранних этапах эмбриогенеза из мезенхимы
вначале образуется соединительнотканный, или перепончатый остов или так
называемые перепончатые кости. Второй этап характеризуется постепенным
замещением соединительнотканной основы хрящевой тканью и образованием хрящевой
кости. Третий этап начинается с образования очагов окостенения и
постепенного вытеснения, хрящевой ткани костной (рис.
57).

2.3.
Кость как орган (костная и хрящевая ткани,
костный мозг, надкостница) и остеогенез
Кость состоит из
органической компоненты, структуры или матрикса, образованного коллагеновыми
тканями и неорганического или минерального компонента, главным образом фосфата
кальция и карбоната кальция, чьи кристаллы состоят из клеток, которые
называются остеобластами. Каждая кость покрыта «оболочкой», которая называется
надкостницей. Внутренний слой надкостницы содержит остеобласты, или
костеобразующие клетки, увеличивающие диаметр кости либо локально, либо по всей
ее длине. Костное вещество вблизи поверхности и надкостница являются плотными,
то есть концентрация костных кристаллов в них большая и в то же время мало
полостей. Эта область кости известна как кортикальный слой кости. В более
глубоких слоях кости находится менее плотное вещество, хотя оно содержит еще
перемычки или трабекулы более плотной кости, проходящей через него. Эта область
называется губчатым веществом кости (рис. 58). (рис. 59)
Как правило, мы не думаем,
что кости нуждаются в снабжении кровью так же, как это делается в коже и
мышцах, но кости во многом являются живыми тканями. Если кости не
обеспечиваются кровью надлежащим образом, то ситуация с костями будет точно
такой же, как и с другими частями тела. Любое состояние, при котором
уменьшается поставка крови к области кости, приводит к боли или хромоте. Кровь
циркулирует через артерии и вены в кость, где находятся так называемые
гаверсовы каналы, которые также содержат лимфатические сосуды и нервы. Большие
длинные кости, такие, как бедренная кость, имеют полости вдоль их центральной
части. Они называются костномозговыми полостями. Эти полости заполнены губчатой
массой из клеток крови, жировых клеток и тканей, которые производят
дополнительно клетки крови.
Поскольку жеребенок растет,
то должны расти и его кости. Это не достигается простым дополнением внешнего
слоя вокруг всей кости, так как это приведет к чрезмерному утолщению костей.
Вместо этого кости имеют специальные области, которые называются пластинами
роста или эпифизом. Эпифиз похож на диск из кости, который закладывает основу
новой кости на каждой стороне и таким образом «выдавливает» конечную часть
кости от центральной оси. Этим способом кость растет в длину. В то же время ее
диаметр увеличивается намного медленнее, что осуществляется в процессе
взросления. Эпифиз является наиболее хрупкой частью скелета. Молодые кости
часто разрушаются вдоль линии такой пластины роста. Неравномерное развитие
отдельных пластин роста порождает деформацию кости и неправильную
пространственную форму. Кроме того, когда пластины роста прекращают свою
деятельность, то в этом случае данная кость прекращает расти.
Давая полную картину
последствий от разрушения кости, необходимо рассмотреть, что будет, если кость
сломается. В этом случае организм лошади пытается противодействовать такому
состоянию путем попыток установить некоторую связь между двумя отдельными
фрагментами. Он делает это с помощью образования волокнистой соединительной
(фиброзной) ткани, которая распространяется от обоих фрагментов кости до тех
пор, пока не произойдет их соединение. Хотя такая соединительная ткань
(состоящая в основном из коллагена) кажется на ощупь твердой, но она не
является достаточно жесткой, чтобы выдерживать напряжение при передвижении.
Поэтому на этом этапе необходимо зафиксировать сломанную кость, чтобы ее
фрагменты еще больше не повредили находящиеся вокруг кровеносные сосуды и
мышцы, но при этом лошадь будет все равно хромать и не сможет использовать ногу
надлежащим образом. В некоторых случаях хотя процесс выздоровления и
заканчивается, но лошадь уже никогда не может полностью использовать ногу, в
которой была сломана кость. Лишь при условии, что волокнистая соединительная
ткань превращается в кость (костенеет), зажившая кость может функционировать
нормально, но в том случае, если между фрагментами сломанной кости есть
какие-либо существенные смещения, то процесс окостенения происходить не будет.
Мацерированная
(обезжиренная, отбеленная и высушенная) кость на 1/3 состоит из неорганических
веществ (оссеин), и на 2/3 из неорганических веществ.
Преобладание в кости раннего
постнатального онтогенеза органических веществ, обеспечивает ей большую
упругость, эластичность.
При изменении соотношения в
сторону преобладания неорганических веществ кость становится ломкой, хрупкой, а
в сторону органических веществ (после обработки соляной кислотой) – мягкой,
гибкой.
Состав свежей кости (По Жеденову В.Н., 1958):Вода – 50%
Коллаген –
12,4%;
Жир – 15,7%;
Минеральные
вещества – 21,9%
Неорганические
вещества:
Ca3(PO4)2 – 85%
CaCO3 – 9.0%
CaF2 – 3.0%
Mg(PO4)2 – 1.5%
NaCL и KCL – 0.5%
Другие – 1,0%
Замещение хрящевой
ткани костной может проходить или изнутри хрящевой кости — энхондральный тнп
окостенения, —оздНсаШ епсюспоп(1-паШ, когда происходит постепенное увеличение
костного ядра, или с ее поверхнасти — перихтдральное окостенение —
оздйсатло репспопагозтаИз, при котором костная ткань нарастает вокруг кости в
виде поверхностного циркулярного костного слоя.
В длинных трубчатых
костях окостенение начинается перихондрально в области диафиза и
распространяется в его глубину, где в толще хряща закладываются очаги
энхондрального окостенения, завершающееся образованием губчатого вещества
кости. После замещения хрящевой ткани костной дальнейшее развитие кости в
толщину происходит за счет остеобластов внутреннего слоя надкостницы, -которая
полностью заменяет бывшую здесь надхрящницу.
Нарастание массы кости с поверхности за счет
надкостницы происходит неравномерно, что сопровождается образованием в
отдельных участках тела кости выемок и бороздок, которые, постепенно
закрываясь, превращаются в канальцы. В силу того что в образовавшихся канальцах
сохраняются остеобласты, то за их счет происходит дальнейшее образование
костного вещества в виде кольцевых пластинок вокруг центрального канала.
Система кольцевых пластинок' вместе с центральным каналом составляет остеон,
являющийся структурной единицей трубчатых костей.
Параллельно с увеличением кости в толщину за
счет особых клеток остеокластов происходят рассасывание (резорбция) ее
губчатого вещества и образование костномозговой полости. В метафизах, где
сохраняется губчатое вещество,, благодаря наличию здесь эндоста и его клеток —
остеобластов осуществляется увеличение кости в длину.
Суставные концы трубчатых костей после начала
окостенения в диафизе еще некоторое время сохраняют хрящевое строение, которое
за счет появления здесь особых энхондральных очагов окостенения постепенно
замещается костной тканью. По мере развития в эпифизах губчатого вещества
происходит его сближение с метафизом и сохранение между ними метаэпифизарной
хрящевой прослойки, которая в силу активного размножения хрящевых клеток долгое
время сохраняется и у взрослых животных. Полное срастание диафиза с метафизом
свидетельствует о завершении роста кости в длину и характеризует наступление
зрелости костяка животного.
Отростки длинных и смешанных костей, на
которых происходит прикрепление Связок и сухожилий мышц, могут иметь свои очаги
окостенения, располагающиеся в глубине хрящевого зачатка, т. е. окостеневают
энхондрально. Энхондральное окостенение присуще и коротким костям.
Вторичные.кости (покровные и многие кости
лицевого отдела черепа) развиваются непосредственно из соединительной ткани
путем закладки в них очагов окостенения (эндесмальное окостенение), от
которых разрост костной ткани происходит в виде постепенно увеличивающихся
лучей (радиальное разрастание костной ткани) до тех пор, пока не произойдет
полное замещение соединительнотканной основы костной. Границы между отдельными
костями, свойственные для черепов молодых животных, постепенно уменьшаются до
полного их исчезновения, что особенно рано происходит в скелете головы птиц.
Таким образом, в зависимости от расположения
очагов окостенения и степени участия в образовании кости остеобластических
источников различают следующие разновидности окостенения: периостальное окостенение,
когда кость развивается из остеогенных клеток надкостницы; эндостальное, если,
в образовании кости участвуют остеобласты внутренней выстилки костномозговой
полости; мезостальное — остеоны образуются из остеобластов
периадвентициального слоя внутрикостных сосудов.
Очаги окостенения закладываются в
определенной последовательности и в определенные сроки онтогенеза, что
обусловлено видом, породой, конституцией животного, а также условиями его
содержания, питания, эксплуатации и физиологическим состоянием.
Сроки зрелости животных (по В. Н. Жеденову с изменениями)
|
Вид животного |
Половая зрелость |
Зрелость тела (завершение дифференциации и роста основных
органов) |
Зрелость костяка (завершение окостенения и роста трубчатых
костей) |
|
Верблюд Лошадь Крупный рогатый скот Мелкий рогатый скот Свинья Собака, кошка Кролик |
2—5 лет 1,5 года 8—12 мес. 7—8 мес.
5—8 мес.
4—8 мес.
4—5 мес.
|
4—5 лет 3 года 2 года 1—1,5 года 9—12 мес. 10-12 мес. 6—8 мес. |
8 лет 4,5—5 лет 4—4,5 года З-3,5 года 3-3,5 года 1,5-2 года 1 год |
Завершение
процесса окостенения у животных свидетельствует о достижении зрелости костяка,
которая не совпадает ни с половой, ни с физической зрелостью тела.
2.4.
Классификация костей по форме, строению,
развитию, структура костной ткани и ее морфофункциональная характеристика
Кости в скелете по форме, строению и по функции
подразделяют на:
- трубчатые: длинные биэпифизарные, длинные
моноэпифизарные, короткие моноэпифизарные;
- короткие ассиметричные;
- плоские;
- смешанные;
- воздухоносные;
- длинные изогнутые.
По происхождению:
- на первичные;
- вторичные (рис. 60).
2.5.
Факторы, влияющие на строение и развитие
костей
· Физическая нагрузка на кость в виде сжатия и растяжения;
· Топографическое расположение и функциональное назначение, направление
силы тяги действующих на них мышц;
· Одновременное протекание двух процессов: разрушение старой костной
ткани (резорбция) и образование новых костных клеток и межклеточного вещества.
2.6.
Осевой скелет
Осевой скелет у изучамых млекопитающих
представлен метамерно расположенными позвонками, формирующими позвоночной
столб, грудной клеткой и черепом (рис. 61).
2.6.1.
Позвоночный столб
В типичном позвоночнике различают 5 отделов: шейный, грудной
(соответствующий грудной клетке), поясничный, крестцовый и хвостовой. Число
шейных позвонков сильно варьирует в зависимости от группы животных. У
современных амфибий такой позвонок только один. У мелких птиц позвонков может
быть всего 5, а у лебедей - до 25. У мезозойской морской рептилии плезиозавра
было 72 шейных позвонка. У млекопитающих их почти всегда 7; исключение
составляют ленивцы (от 6 до 9). У китообразных и ламантинов шейные позвонки
частично слиты и укорочены в соответствии с укорочением шеи (по мнению ряда
специалистов, у ламантинов их всего 6). Первый шейный позвонок называется
атлантом. У млекопитающих и амфибий на нем две суставные поверхности, в которые
входят затылочные мыщелки. У млекопитающих второй шейный позвонок (эпистрофей)
образует ось, на которой вращаются атлант и череп. К грудным позвонкам обычно
присоединены ребра. У птиц их около пяти, у млекопитающих 12 или 13; у змей
очень много. Тела этих позвонков обычно малы, а остистые отростки их верхних
дуг длинные и наклонены назад. Поясничных позвонков обычно от 5 до 8; у
большинства рептилий и всех птиц и млекопитающих они не несут ребер. Остистые и
поперечные отростки поясничных позвонков очень мощные и, как правило,
направлены вперед. У змей и многих рыб ребра причленены ко всем туловищным
позвонкам, и границу между грудным и поясничным отделами провести трудно. У
птиц поясничные позвонки слиты с крестцовыми, образуя сложный крестец, что
делает их спину более жесткой, чем у других позвоночных, исключая черепах, у которых
грудной, поясничный и крестцовый отделы соединены с панцирем. Число крестцовых
позвонков варьирует от одного у амфибий до 13 у птиц. Строение хвостового
отдела тоже весьма разнообразно; у лягушек, птиц, человекообразных обезьян и
человека он содержит всего несколько частично или полностью слившихся
позвонков, а у некоторых акул - до двухсот. Ближе к концу хвоста позвонки
утрачивают дуги и представлены одними телами.
2.6.1.1.
Шейный отдел
Шейный
отдел – pars
cervicales – у всех млекопитающих представлен семью позвонками,
исключение ленивец (6 позвонков): третий, четвертый, пятый и шестой – типичные
позвонки, первый, второй и седьмой – атипичные и имеют отличительные
особенности в строении от типичного позвонка.
Типичный
позвонок – vertebra.
Строение:
Тело позвонка – corpus veretebrae.
На теле: краниально –
головка позвонка – caput vertebrae,
каудально – ямка позвонка – fossa
vertebrae, вентрально – вентральный гребень – crista ventralis, латерально – поперечные
отростки – processus
transverzarius, разделенные на две лопасти и боковые позвоночные
отверстия – foramen vertebralis
lateralis.
Над телом – дужка позвонка –
arcus vertebrae.
На дужке: краниально и
каудально – парные суставные отростки – processus articularis cranialis et caudalis,
дорсально – один непарный остистый отросток – processus spinosus.
Между дужкой и телом
краниально и каудально – вырезки – incissura cranialis et caudalis, которые при соединении
двух смежных позвонков образуют межпозвоночные отверстия – foramen intervertebralis (рис. 62).
Первый
шейный позвонок – атлант – atlant, atlas, обеспечивает подвижность головы и
имеет следующие отличительные особенности в строении от типичного позвонка:
- не имеет тела;
- имеет две дужки –
дорсальную и вентральную;
- два бугорка: дорсальный и
вентральный – tuberculum dorsalis
et ventralis;
- сросшиеся поперечные
отростки образуют крылья атланта – ala atlantis, вентрально на крыльях – крыловые ямки – fossa alaris. В ямках – крыловые
отверстия, которые соединяются желобком с межпозвоночными отверстиями – foramen alaris et intervertebralis;
- суставные поверхности:
краниальная для мыщелков затылочной кости и каудальная для второго шейного
позвонка (рис. 63).
Второй
шейный позвонок – epystrophey, axis:
- мощный дорсальный гребень
– crista dorsalis;
- отсутствие краниальных
суставных отростков и как следствие, наличие межпозвоночных отверстий – foramen intervertebralis;
- краниально вместо головки
– наличие зубовидного выступа – dens
epystrophey;
- поперечно-реберные
отростки не разделены на две лопасти (рис. 64).
Седьмой
шейный позвонок:
- наличие реберных фасеток –
fovea costalis – у ямки позвонка;
- более мощный остистый
отросток;
- отсутствие боковых
позвоночных отверстий;
- поперечно-реберные
отростки не разделены на две лопасти (рис. 65).
Видовые особенности:
Лошадь –
типичные позвонки очень массивные и длинные, головки и ямки хорошо выражены,
остистые отростки отсутствуют, вместо них шероховатости. Вентральные гребни
хорошо выражены.
Крылья атланта с округлыми
краями, сильно загнуты вентрально, крыловые ямки глубокие,
Эпистрофей самый мощный и
длинный из всех позвонков. Зубовидный выступ с плоской дорсальной поверхностью.
Дорсальный гребень каудально раздваивается и срастается с каудальными
суставными отростками. Вентральный гребень ярко выражен, ямка позвонка
глубокая.
Седьмой шейный позвонок
имеет относительно небольшой остистый отросток.
Крупный
рогатый скот – типичные позвонки массивные, короткие,
головки и ямки хорошо выражены, остистые отростки развиты, имеют утолщенные
концы. Вентрального гребня нет.
Крылья атланта мощные.
Крыловая ямка мелкая.
Зубовидный выступ эпистрофея
полуцилиндрической формы, дорсальный гребень имеет почти квадратную форму.
У седьмого шейного позвонка
ярко выражен остистый отросток, вентрального гребня нет.
Свинья –
типичные позвонки массивные, но очень короткие, головки и ямки плоские, дужки
узкие. Остистые отростки узкие и длинные. Вентрального гребня нет.
Крылья атланта узкие и
массивные. У основания крыльев начинается поперечный канал, идущий в крыловую
ямку. Дужки позвонка узкие. Вентральный бугорок узкий и выступает
вентрокаудально.
Зубовидный выступ эпистрофея
конусовидной формы, дорсальный гребень массивны и высокий. Тело позвонка
короткое, ямка плоская.
У седьмого шейного позвонка
головка и ямка плоские, помимо межпозвоночного отверстия имеется латеральное
отверстие
Собака – у
типичных позвонков головка и ямка плоские. Дуги позвонков широкие, остистый
отросток на 3-м позвонке отсутствует.
Крылья атланта плоские,
широко расставлены, дорсальная дужка широкая, не имеет дорсального бугорка.
Вместо крылового отверстия – крыловая вырезка.
Второй позвонок длинный,
имеет широкий гребень, который нависает над зубовидным выступом. Каудальный
край срастается с каудальными суставными отростками. Межпозвоночных отверстий
нет, вместо них – вырезки.
На седьмом шейном позвонке
остистый отросток узкий и высокий
2.6.1.2.
Грудной отдел
Грудная клетка – thorax – представляет собой конус с усеченной
вершиной и широким косо поставленным основанием. Имеется вход в грудную полость
(вершина конуса) и выход (основание конуса) – apertura craniales et caudales.
Грудная клетка состоит из грудных позвонков, ребер и грудины.
Грудной позвонок – vertebra thoracicae – имеет следующие
отличительные особенности от типичного позвонка:
- остистые отростки ярко
выражены, имеют неодинаковую величину (наиболее высокие в области холки) и
каудальный наклон, имеется позвонок с вертикально стоящим остистым отростком –
это диафрагмальный позвонок (на его уровне прикрепляется диафрагма);
- наличие реберных фасеток –
fovea costalis – у
головки и у ямки позвонка;
- отсутствие суставных
отростков и наличие суставных фасеток – fovea articularis, исключение – последние
грудные позвонки;
- поперечные отростки
расположены не на теле, а на дужке позвонка, имеют реберную фасетку для
прикрепления бугорка ребра и сосцевидный отросток – processus mammilaris (рис.
66).
Ребро – costa – состоит из длинной изогнутой кости (os costae) и реберного хряща (cartilage costae).
На дорсальном конце ребра:
- головка ребра – capitulum costae, с двумя реберными
фасетками для прикрепления к двум смежным позвонкам;
- бугорок ребра – tuberculum costae, с одной реберной фасеткой
для прикрепления к поперечному отростку грудного позвонка;
- шейка ребра – collum costae.
Ниже бугорка ребра находится
угол ребра – angulus costae, он
придает объем грудной клетке.
На теле ребра:
- латерально – мышечный
желоб (sulcus muscularis)
для прикрепления межреберных мышц;
- медиально – сосудистый
желоб (sulcus vascularis)
для прохождения межреберных сосудов.
Посредством реберных хрящей ребра
соединяются с грудиной – истинные ребра (стернальные) или образуют реберную
дугу – ложные ребра (астернальные) (рис. 67).
Грудная
кость – sternum –
образует вентральную стенку грудной клетки, она состоит из рукоятки, тел и
мечевидного отростка.
Рукоятка – manubrium sterni – лежит впереди
прикрепления второй пары ребер;
Тела – sternebra, это костные сегменты,
соединенные в зависимости от возраста, хрящом или костной тканью. Латерально на
каждом сегменте располагаются реберные вырезки для прикрепления реберных
хрящей;
Мечевидный отросток – processus xyphoideus, заканчивается
мечевидным хрящом – cartilago xyphoideus (рис. 68).
Видовые особенности:
Лошадь
– 18-19
грудных позвонков, вентральные гребни хорошо развиты. Диафрагмальный позвонок
15-й.
18-19 пар ребер, их ширина
одинакова на все протяжении. Стернальных ребер 8 пар.
Грудина сжата с боков,
килевидной формы. Рукоятка срастается с телом, краниально имеется хрящ
(соколок), который формирует вентральный гребень. Мечевидного отростка нет,
есть только мечевидный хрящ.
Крупный
рогатый скот – 13 грудных позвонков. Остистые отростки
широкие и пластинчатые. Диафрагмальный – 13-й. имеется самостоятельное
межпозвоночное отверстие.
13 пар ребер. Ширина ребер
неравномерная – дорсальный конец в 2-3 раза уже грудинного. Стернальных 8 пар.
Кость сжата в
дорсовентральном направлении. Рукоятка массивная, соединяется с телом суставом
Свинья –
14-15 грудных позвонков, их тела короткие с плоскими головками и ямками,
вентральные гребни отсутствуют. Диафрагмальный позвонок – 11-й. Имеется два
отверстия: латеральное и межпозвоночное.
14-15 пар ребер. 7 пар –
истинные. Ребра узкие, равномерно округлые, сильно изогнуты, шейка ребра длинная.
Рукоятка грудины массивная,
латерально сжата. Тело состоит из 6 сегментов, сжато дорсовентрально,
расширяется в каудальном направлении. Мечевидный отросток удлиненный.
Собака –
13 грудных позвонков. Диафрагмальный – 11-й. На последних позвонках – добавочные
отростки.
13 пар ребер. 9 пар –
истинные. Ребра узкие, равномерно округлые.
Грудная кость состоит из 8
сегментов. Рукоятка и мечевидный отросток имеют большую длину. Мечевидный хрящ
небольшой.
2.6.1.3.
Поясничный отдел
Поясничный позвонок – vertebra lumbales, имеет следующие
отличительные особенности от типичного позвонка:
- поперечные отростки имеют пластинчатую форму и большую
величину;
- остистые отростки имеют одинаковую высоту и краниальный
наклон;
- головки и ямки – плоские;
- наличие сосцевидные отростков на краниальных суставных
отростках (рис. 69).
Видовые
особенности:
Лошадь
– 5-6
поясничных позвонков, суставные отростки с плоскими фасетками. Поперечнореберные
отростки поставлены горизонтально. Высота стистых отростков превышает их ширину
Крупный
рогатый скот – 6 позвонков. Поперечноребрные отростки слегка
изогнуты краниально. Высота и ширина остистых отростков примерно одинакова.
Каудальные межпозвоночные вырезки глубокие.
Свинья – 7
позвонков, их тела короткие. Поперечнореберные отростки дугообразно изогнуты
вентрально.
Собака – 7
позвонков их тела длинные, вентральные гребни отсутствуют. Остистые отростки,
узкие, расстояние между отростками равно их ширине. Поперечноребрные отростки
узкие и направлены вентрокраниально.
2.6.1.4.
Крестцовый отдел
Крестцовая кость – os sacrum, состоит из сросшихся
крестцовых позвонков.
На вентральной поверхности крестцовой кости:
- поперечные линии – liniae transversae – место сращения позвонков;
- межпозвоночные отверстия.
На дорсальной поверхности:
- остистые отростки, в зависимости от вида животного,
срастаются в основании, отсутствуют совсем или образуют дорсальный гребень;
- суставные отростки при сращении образуют латеральные
гребни;
- поперечные отростки при сращении образуют боковые части
– pars lateralis;
- межпозвоночные отверстия;
- на первых двух позвонках поперечные отростки образовали
крылья крестцовой кости – ala
sacrales,
на латеральной поверхности которых имеются ушковидные суставные поверхности – facies articularis auricularis;
- передний вентральный край первого позвонка называется
мыс – promontorium (рис. 70).
Видовые
особенности:
Лошадь
– 5
позвонков образуют кость. Остистые отростки срастаются только в основании.
Крупный
рогатый скот – 5 позвонков. Остистые отростки образуют
мощный гребень с утолщением по дорсальному краю. Крылья кости сжаты
краниокаудально.
Свинья –
кость образую 4 позвонка. Остистые отростки отсутствуют ушковидные суставные
поверхности обращены латерально.
Собака – 3
позвонка. Остистые отростки срастаются только в основании.
2.6.1.5.
Хвостовой отдел
- остистые, суставные, поперечные отростки и дужки
сохраняются на первых пяти позвонках, соответственно, здесь сохраняется
позвоночный канал;
- головки и ямки выпуклые, которые обеспечивают
наибольшую подвижность в этом отделе позвоночного столба;
- тела в каудальном направлении укорачиваются и
истончаются, отростки постепенно превращаются в бугорки, а затем полностью исчезают
(рис. 71).
Видовые особенности:
Лошадь – хвост относительно короткий, позвонки в количестве 15-20,
очень короткие, массивные, цилиндрические. Дужки выражены на первых трех
позвонках, остистых отростков нет.
Крупный
рогатый скот – хвост длинный, в количестве 18-20, их тела
вытянутые и длинные. Под телами первых пяти позвонков имеются гемальные дужки –
arcus haemales,
для прохождения срединной хвостовой артерии.
Свинья – позвонки
длинные, в количестве 20-23, на первых пяти позвонках сохраняются дужки,
поперечные отростки широкие
Собака
- 20-23 хвостовых позвонков, на первых
пяти позвонках имеются гемальные дужки.
2.6.2.
Скелет головы – череп животных
На переднем конце биполярного
тела животного развивается два самостоятельных раздела скелета головы. Один –
защитное приспособление для головного мозга (мозговой отдел), второй (лицевой
отдел) – для лучшего, большего захвата пищи и ее первичной обработки (рис. 72).
2.6.2.1.
Затылочная и клиновидная кости черепа
Затылочная
кость – os
occipitale – непарная, состоит из тела, двух боковых частей и чешуи,
между этими структурами находится большое затылочное отверстие – foramen occipital magnum (это
граница соединения продолговатого и спинного мозга).
Тело – pars
basilaris – находится на бальной поверхности черепа и формирует
его основание. Латерально у лошади и свиньи имеются рваные отверстия – foramen lacerum. На границе с телом
клиновидной кости имеются мышечные бугорки или отростки для прикрепления
длинных мышц головы. На мозговой поверхности тела имеются ямки для
продолговатого мозга (fossa
medullae oblongatae) и мозгового моста (fossa pontis).
На боковых частях – pars laterals – различают мыщелки – condylus occipitalae, для сочленения с
атлантом. Латерально от мыщелков находятся яремные отростки – processus jugularis. Между мыщелком и яремным
отростком находится подъязычное отверстие – foramen hypoglossi, для выхода подъязычного
нерва.
Чешуя затылочной кости – squama occipitalis, имеет выйную поверхность –
facies nuchalis, с
затылочным гребнем. На поверхности
имеется выйное возвышение или затылочное предбугорье – protuberantia occipitalis –
для прикрепления выйной связки. На внутренней поверхности чешуи имеются пальцевые
вдавления (отпечаток мозжечка) – impressionale
digitalis (рис. 73).
Клиновидная
кость – os
sphenoidale – непарная, располагается в основании черепа, состоит из
тела, двух пар крыльев и крыловидных отростков.
Тело – corpus
sphenoidale, образовано двумя частями: базисфеноидом и пресфеноидом.
На мозговой поверхности тела имеется ямка турецкого седла – fossa sella turcica, для гипофиза и желоб
зрительного перекреста для зрительных нервов.
Латерально от базисфеноида располагаются височные крылья
– ala temporalis, а
от пресфеноида – глазничные крылья – ala orbitalis. На границе крыльев имеется крыловидный
гребень – crista pterygoideus, у
основания которого четыре отверстия:
решетчатое (foramen ethmoidale),
зрительное (foramen opticum),
глазничная щель (fissura
orbitalis), круглое отверстие (foramen rotundum).
Вентрально от тела отходят крыловидные отростки, в
основании которых имеется крыловые каналы (рис. 74).
Видовые
особенности:
Лошадь
–
в затылочной кости хорошо выражено рваное отверстие, яремные отростки прямые,
вентрально от выйного возвышения – выйная ямка. Затылочный гребень высокий.
В клиновидной кости присутствуют все четыре отверстия. У
основания крыловидного отростка имеется крыловой канал.
Крупный
рогатый скот – яремные отростки короткие и загнуты
медиально, два парных подъязычных отверстия. Затылочный гребень отсутствует.
Рваное отверстие щелевидное.
На границе крыльев клиновидной кости три отверстия:
решетчатое может располагаться на лобной кости, а глазничная щель
Свинья –
яремные отростки длинные и прямые, чешуя высокая, отсутствует выйное
возвышение. Рваное отверстие слабо выражено.
Крыловидные отростки большие, крыловой гребень высокий.
Имеются круглоглазничное отверстие и две вырезки – овальная и сонная.
Собака –
яремные отростки короткие и прямые. Имеется хорошо выраженный затылочный
гребень. Рваное отверстие лежит позади барабанного пузыря височной кости.
Височные крылья клиновидной кости выражены сильнее, чем
глазничные. Имеются крыловое и овальное отверстия. Глазничная щель и круглое
отверстие образуют круглоглазничное отверстие.
2.6.2.2.
Теменная, межтеменная, лобная, височная,
крыловидная кости черепа
Теменная
кость – os
parietale – парная пластинчатая кость, участвует в образовании височной
ямки. На мозговой поверхности имеются пальцевые вдавления, мозговые гребешки – juga cerebellum и сосудистые желоба.
Межтеменная
кость - os
interparietale – непарная, треугольной или квадратной формы,
выражена только у плодов и новорожденных.
Лобная
кость – os
frontale – парная, пластинчатая кость. Состоит из пластинчатой
части, глазничной и носовой. Пластинчатая часть имеет надглазничные отверстия (foramen supraorbitalis) и скуловой отросток
(processus zygomaticus).
Глазничная часть участвует в образовании медиальной стенки глазницы. Носовая
часть участвует в образовании крыши носовой полости. Лобная кость содержит в
себе лобную пазуху (sinus frontalis).
Височная кость – os temporale - парная, образует
латеральные стенки мозговой полости. В ней расположен вестибулярно-слуховой
орган. Кость граничит с затылочной, клиновидной и теменными костями.
Различают
чешую височной кости и каменистую кость. Чешуя – squama temporale – довольно обширна,
принимает участие в формировании височной ямки и несет скуловой отросток.
Последний имеет суставную поверхность для сочленения с нижней челюстью и
засуставной отросток. Скуловой отросток участвует в образовании скуловой дуги.
Латеральный край имеет височный гребень. У основания скулового отростка
расположено височное отверстие, а между наружным слуховым проходом и
засуставным отростком – височный канал для мозговой вены.
Каменистая кость – os petrosum – состоит из барабанной,
сосцевидной и скалистой частей. Барабанная часть лежит между скуловым отростком
височной кости и телом затылочной кости. В ее состав входит наружный слуховой
проход длиной 1,8-
Скалистая
часть несет внутренний слуховой проход, на дне которого имеется переднее
отверстие – для лицевого нерва и заднее – для VIII пары (равновесно-слуховой нерв).
Скалистый гребень делит мозговую полость на переднюю ямку для большого мозга и
заднюю для ромбовидного. Дорсально от внутреннего слухового прохода расположено
отверстие водопровода преддверия, а несколько ниже – узкая щель водопровода
улитки.
Сосцевидная
часть представлена одноименным отростком и узкой полоской, которая вклинивается
между чешуей затылочной и височной костями. Между наружным слуховым проходом и
сосцевидным отростком имеется отверстие лицевого канала.
Крыловидная
кость – os
pterygoideum, парная, прилегающая с медиальной стороны к вентральной
пластинке небной кости и крыловидному отростку клиновидной кости. Нижний конец
ее заканчивается слабо выраженным крючком (рис. 75).
Видовые
особенности:
Лошадь
–
на наружной поверхности теменной кости – сагиттальный гребень, который назально
переходит в наружный лобный гребень. Межтеменная кость имеет четырехугольную
форму.
Скуловой отросток лобной кости соединяется со скуловым
отростком височной кости. В основании отростка – надглазничное отверстие. На
орбитальной поверхности находится блоковая ямка – fossa throchlearis. По краям ямки
прикрепляется хрящевая пластинка через которую перекидывается сухожилие
дорсальной косой мышцы глаза.
Каменистая и барабанная части височной кости срастаются
только к старости. Наружный слуховой проход короткий. Хорошо развит мышечный
отросток.
Крыловидная кость узкая и длинная, крючок слабо изогнут.
Крупный
рогатый скот – теменные кости делятся на две части:
затылочную и височную, которые разделены височным гребнем. Межтеменная кость
имеет треугольную форму.
Лобные кости развиты сильно. Имеются межроговый гребень и
мощные роговые отростки. Скуловой отросток короткий и соединяется со скуловым
отростком височной кости. Надглазничное отверстие двойное, от него отходит
надглазничный желоб. В самой кости и роговом отростке имеются лобные пазухи.
Суставной бугорок височной кости массивный, на нем
выделяется подъязычный отросток. Наружный слуховой проход длинный.
Крыловидная кость длинная и широкая крючок притуплен.
Свинья –
теменная кость массивная, ее наружная поверхность разделена гребнем.
Межтеменная кость отсутствует.
Наружная поверхность лобной может плоской или вогнутой в
зависимости от породы. Надглазничное отверстие открывается на уровне переднего
края орбиты. Скуловой отросток короткий, орбита не замкнута.
Скуловой отросток височной кости массивный и короткий,
наружный слуховой проход длинный. Барабанный пузырь вытянутой формы.
Крыловидная кость имеет крыловидную ямку. Крючок
притуплен.
Собака –
на теменной кости выражен сагиттальный гребень. Межтеменная кость сердцевидной
формы.
Скуловой отросток лобной кости развит слабо. Надглазничное
отверстие отсутствует.
Наружный слуховой проход височной кости короткий,
барабанный пузырь округлый.
Крыловидная кость короткая, широкая, с коротким крючком.
2.6.2.3.
Носовая, слезная, решетчатая, скуловая,
резцовая, небная кости черепа, носовые раковины
Парная
носовая
кость – os nasale –
образует дорсальную стенку носа, граничит с лобной, верхнечелюстной и резцовой
костями, имеет форму треугольника. На медиальной поверхности ее расположен
дорсальный раковинный гребень.
Впереди
кость заканчивается ростральным раздвоенным отростком, между которым и носовым
отростком резцовой кости образуется носорезцовая вырезка.
Слезная
кость – os
lacrimale – парная, граничит с лобной, носовой, верхнечелюстной и
скуловой костями и состоит из лицевой и орбитальной частей. На границе
пластинок, по краю орбитального гребня расположено слезное отверстие, ведущие в
слезно-носовой канал. На лицевой пластинке имеется предглазничная ямка, а на
орбитальной пластинке хорошо выражена ямка для вентрального косого мускула
глаза.
Решетчатая
кость – os
ethmoidale, непарная, лежит на границе носовой и мозговой полости.
Состоит из решетчатой, перпендикулярной, боковой пластинок и лабиринта.
Решетчатая
пластинка – lamina ethmoidale –
имеет многочисленные отверстия для обонятельного нерва и формирует две
обонятельные ямки, разделенные петушьим гребнем. В обонятельные ямки
открывается решетчатое отверстие.
Лабиринт кости – labirintus ethmoidale – состоит из завитков
тонких костных пластинок – турбиналий, из которых две наиболее крупные вдаются
в носовую полость.
Перпендикулярная пластинка – lamina perpendicularis – является
продолжением петушьего гребня и рострально переходит в носовую перегородку.
Боковые пластинки оформляют наружные стенки лабиринта (рис. 76).
Скуловая
кость – os
zygomaticum – парная, граничит со слезной, лобной, верхнечелюстной и
скуловым отростком височной кости, принимает участие в формировании вентральной
части орбиты и скуловой дуги. Различают тело скуловой кости и два отростка:
лобный и височный. Первый направлен в сторону скулового отростка лобной кости,
второй – к височной кости и образует скуловую дугу. На теле кости имеется
скуловой гребень, под которым закрепляется m. masseter, а впереди между верхнечелюстной
и небной костями лежит в виде узкого треугольника слезно-челюстной отросток.
Резцовая кость – os incisivum, парная, расположена
впереди от верхнечелюстной кости и состоит из тела, носового и небного
отростков. Тело короткое, не имеет резцовых луночек. Носовой отросток направлен
к одноименной кости и формирует костный
вход в носовую полость. Небный отросток образует вместе с носовым отростком
небную щель.
Небная
кость – os
palatinum, парная, расположена позади твердого неба, принимает
участие в формировании хоан и состоит из горизонтальной и вертикальной
пластинок. Горизонтальная пластинка рострально соединяется с небными
пластинками верхней челюсти и на носовой поверхности несет гребень для сошника.
Между горизонтальной пластинкой и небным отростком верхней челюсти проходит
небный канал, который впереди открывается большим, а сзади малым небным
отверстием.
Вентральная
или перпендикулярная пластинка участвует в образовании крылонебной ямки и
ограничивает с боковых сторон хоаны. В передней ее части имеется клинонебное
отверстие.
Носовые раковины – conchae nasalis – парные, расположены на латеральной поверхности носовой полости. Это
пористые костные пластины, закрученные веретенообразно, служат для увеличения
площади слизистой оболочки носа и образования носовых ходов.
Дорсальная раковина
прикрепляется к раковинному гребню – crista conchalis – носовой кости и решетчатой кости.
Вентральная носовая
раковина прикрепляется к раковинному гребню верхнечелюстной кости.
Видовые особенности:
Лошадь
–
назальный конец носовой кости заострен хорошо выражен раковинный гребень.
Размеры лицевой и глазничной пластинок слезной кости
почти одинаковы.
В решетчатой кости 6 эндотурбиналий и 25 эктотурбиналий.
Лицевая поверхность скуловой коти обширная, на ней есть
скуловой гребень, переходящий в лицевой.
Тело резцовой кости массивное с тремя альвеолами для
резцов. Между костями – резцовый канал.
Горизонтальная пластина небной кости узкая
перпендикулярная соединяется с клиновидной костью.
Дорсальная носовая раковина сильно развита.
Крупный
рогатый скот – назальный конец носовой кости раздвоен,
хорошо выражен раковинный гребень.
Лицевая пластинка слезной кости длинная и достигает
носовой кости. У взрослых животных имеется слезный пузырь, который сообщается с
верхнечелюстной пазухой.
В лабиринте решетчатой кости 5 эндотурбиналий и 18
эктотурбиналий.
Скуловая кость имеет два отростка: скуловой и лобный. На
лицевой поверхности выражен лицевой гребень.
Тело резцовой кости плоское в виде пластинки из-за
отсутствия резцов, между костями – межрезцовая щель.
Горизонтальная пластинка небной кости широкая и участвует
в образовании небной пазухи.
Вентральная носовая раковина короче и шире дорсальной.
Свинья –
носовая кость узкая и длинная, на ее поверхности проходит надглазничный желоб.
Слезное отверстие двойное и открывается на орбитальном
крае. На лицевой пластинке имеется слезный бугорок. С верхнечелюстной кости
продолжается клыковая ямка.
В решетчатой кости 7 эндо- и 20 эктотурбиналий.
Лицевая пластинка скуловой кости хорошо развита. Имеется
небольшой лобный отросток, орбита не замкнута. Височный отросток высокий.
На теле резцовой кости находятся три далеко расставленные
друг от друга альвеолы для резцов. Носовой отросток широкий и массивный.
Горизонтальная пластинка небной кости сильно развита,
перпендикудярная – слабая.
Дорсальная носовая раковина слабо развита, костные
пластинки вентральной раковины достаточно прочные.
Собака
– конец носовой кости сильно расширен и
вырезкой разделен на два отростка.
На слезной кости развита только глазничная пластинка.
В решетчатой кости 4 эндотурбиналии и 6 эктотурбиналии.
Скуловая кость несет четыре отростка: слезный,
верхнечелюстной, лобный и височный.
Тело резцовой кости плоское, носовой отросток длинный,
небный отросток сжат с боков.
Горизонтальная пластина небной кости широкая.
Вентральная носовая раковина, образуя сложный комплекс
костных трубочек.
2.6.2.4.
Сошник, верхнечелюстная, нижнечелюстная,
подъязычная кости черепа
Сошник – vomer, непарная кость, располагается от
пресфеноида, где образует крылья, далее следует по сагиттальной линии дна
носовой полости рострально. Верхний край сошника несет желоб для хрящевой
носовой прегородки.
Верхнечелюстная кость – os maxilla, парная, оформляет боковые
стенки носовой и ротовой полостей. Граничит с носовыми, резцовыми, слезными,
скуловыми, небными костями. Тело кости образовано наружной – лицевой и
внутренней – носовой пластинками, соединяющимися в области альвеолярного края.
Последний несет шесть луночек для коренных зубов и аборально заканчивается
верхнечелюстным бугром, с медиальной стороны которого открывается
верхнечелюстное отверстие, ведущее в подглазничный канал. Впереди кость имеет
беззубый край, заканчивающийся луночкой для клыка. Над четвертым коренным зубом
имеется слабо выраженный лицевой бугор, а над передним краем первого премоляра
расположено подглазничное отверстие.
С
нижней стороны, между зубными краями располагаются небные пластинчатые
отростки, которые соединяясь по сагиттальной линии, образуют твердое небо,
граничащее аборально с горизонтальной пластинкой небной кости. Здесь же
открывается небный канал своим передним отверстием – foramen palatinus majus, который
продолжается в небольшой небный желоб. В толще небных отростков находится
небная пазуха.
Носовая
пластинка верхнечелюстной кости образует боковую стенку носовой полости, по
медиальной поверхности которой проходит вентральный раковинный гребень. На
уровне третьего премоляра, дорсально от раковинного гребня имеется отверстие в
верхнечелюстной синус и проходит слезно-носовой канал, открывающийся на уровне
первого премоляра в носовую полость.
На
границе с глазничным крылом клиновидной кости кость образует клинонебную ямку,
в которой дорсовентрально располагаются три отверстия:
Верхнечелюстное – foramen maxillare – это начало подглазничного
канала – canalis
infraorbitalis, который заканчивается подглазничным отверстием – foramen infraorbitale;
Клинонебное – foramen sphenopalatinum – образует канал,
который идет в носовую полость;
Каудальное небное отверстие – foramen palatinum caudale –
является началом небного канала, который заканчивается большим небным
отверстием – foramen palatinum
major –
на горизонтальной пластинке небной кости или на границе верхнечелюстной кости и
небной.
Нижняя
челюсть – mandibula, парная, состоит из рецовой, молярно частей
и двух ветвей. На резцовой части справа и слева имеются луночки для резцов. Обе
половины кости соединяются швом или синостозом. На коренной части расположены
луночки для коренных зубов. Между резцовыми и коренными зубами имеется беззубый
край. Вентральный край кости несет пологую сосудистую вырезку.
Ветвь
– ramus mandibularis,
заканчивается венечным (мышечным) и суставным отростками, между которыми
имеется нижнечелюстная вырезка. Венечный отросток является местом крепления
височного мускула. Суставной отросток служит для соединения со скуловым
отростком височной кости.
Латеральная
поверхность крыла несет шероховатую ямку для большого жевательного мускула, а
медиальная поверхность – ямку крылового мускула. На дне последней имеется
нижнечелюстное отверстие, которое ведет в нижнечелюстной канал. На резцовой
части он открывается подбородочным отверстием (рис. 77).
Подъязычная
кость – os
hyoideum,
непарная, служит местом крепления мускулатуры глотки и языка, а также опорой
гортани. Она состоит из непарного тела и двух парных ветвей. Различают
гортанные (нижние) рога, соединяющиеся со щитовидным хрящом гортани, и малые
рога, направленные дорсально. К последним крепятся ветви, состоящие из
проксимального, среднего и дистального члеников. Проксимальный членик
закрепляется на подъязычном отростке каменистой кости. Средний членик, самый
большой, несет проксимальный и дистальный мышечные бугорки, а дистальный членик
соединяется с малыми рогами (рис. 78).
Видовые
особенности:
Лошадь – сошник соединяется с костным небом, начиная с
хоанного края небных костей.
Верхнечелюстная кость массивная, имеет на
лицевой поверхности мощный лицевой гребень – crista facialis. Хорошо выражен беззубый край, у самцов
имеется альвеола для клыка. Верхнечелюстная пазуха обширная, делится на
переднюю и заднюю камеры.
Нижнечелюстные кости рано срастаются между
собой. На беззубом крае у самцов альвеолы для клыка. Мышечный отросток узкий и
длинный.
Язычный отросток на теле длинный и сжат с
боков, большие рога соединяются с телом с помощью костной ткани, дистальный
членик имеет вид чечевицеобразной косточки, средний членик имеет
шилоподъязычный угол.
Крупный
рогатый скот – сошник к небным костям не прикрепляется, а
ростральный конец сошника не достигает тела резцовой кости.
На лицевой поверхности верхнечелюстной кости находится
лицевой бугор. Верхнечелюстная пазуха обширная, каудально преходит в пазуху
слезного пузыря. В толще небного отростка расположена небная пазуха.
Межальвеолярный край обширный.
На альвеолярном крае нижнечелюстной кости расположены
четыре луночки для резцов, альвеол для клыков нет, шесть луночек для коренных
зубов Мышечный отросток длинный, уставной отросток седлообразный.
Язычный отросток подъязычной кости короткий большие рога
с телом имеют костное сращение, дистальный членик в виде цилиндра.
Свинья –
сошник ростральным концом заканчивается на теле резцовой кости.
На верхнечелюстной кости семь луночек для коренных зубов,
обширная луночка для клыка. На лицевой поверхности выражена клыковя ямка – fossa caninus.
Нижнечелюстные кости сращены в непарную кость. Луночек
для коренных зубов семь, межальвеолярный край слабо выражен. Подбородочных
отверстий несколько. Мышечный отросток узкий и короткий.
На теле подъязычной кости нет язычного отростка, большие
рога с телом соединяются хрящом, дистальный членик имеет вид связки, в верхней
части состоит из хряща.
Собака –
тело сошника не прикрепляется к небным костям.
Верхнечелюстной бугор слабо развит, большое небное
отверстие открывается на границе с небной костью, челюстная пазуха слабо
развита.
Вентральный край нижней челюсти выпуклый. На челюстном
углу – угловой отросток. Ямка для большого жевательного мускула глубокая.
Мышечный отросток широкий и высокий. Луночек для коренных зубов семь, беззубый
край отсутствует.
На теле подъязычной кости нет язычного отростка, большие
рога соединяются с телом с помощью сустава, дистальный членик в форме цилиндра.
2.7.
Периферический скелет и его звенья
2.7.1.
Скелет плечевого пояса и передней конечности
2.7.1.1.
Лопатка, плечевая кость
Лопатка – scapula – представляет плоскую кость
треугольной формы, основание которой направлено дорсо-каудально, а вершина – кранио-вентрально.
Такое расположение лопатки обусловлено действием мышц, прикрепляющихся к кости.
На
основании лопатки имеется надлопаточный хрящ. Суставной угол лопатки несет
суставную поверхность для соединения с головкой плечевой кости и бугор лопатки для
крепления сухожилия двуглавого мускула. На медиальной поверхности бугра
расположен коракоидный отросток для одноименного мускула. Суженная часть
лопатки называется шейкой.
Латеральная
поверхность разделена остью, заканчивающейся акромионом на две ямки –
предостную и заостную. На середине ости имеется небольшой шероховатый бугорок.
Обе ямки являются местами расположения одноименных мускулов.
На
медиальной поверхности лопатки различают подлопаточную ямку для одноименного
мускула и зубчатую шероховатость для вентрального зубчатого мускула.
Краниальный
край лопатки слегка выгнут, а каудальный более прямой (рис.
79).
Скелет
свободной грудной конечности имеет трехзвеньевое строение.
Верхнее звено – стилоподий – включает плечевую кость, среднее звено -
зейгоподий – состоит из лучевой и локтевой костей, третье звено – автоподий
имеет в своем составе кости запястья (базиподий), пясти (метаподий) и пальцев
(акроподий).
Плечевая
кость – os
humeri
(brachii) –
представляет длинную трубчатую кость, на проксимальном конце имеющую головку и
два мышечных бугра – латеральный и медиальный, между которыми расположен желоб
для сухожилия двуглавого мускула. Латеральный (или большой) и медиальный (или
малый) бугры – tuber lateralis
et medialis – служат местами крепления предостного мускула. Головка
переходит в шейку кости. От латерального бугра вниз на диафиз кости опускается
гребень с дельтовидной шероховатостью на нем. На медиальной поверхности тела
кости имеется круглая шероховатость (верхняя треть).
Дистальный
эпифиз несет суставной блок для головки лучевой кости и два мыщелка,
разделенные локтевой ямкой. Латеральный мыщелок отогнут в сторону, медиальный
направлен вертикально. На этих образованиях имеются надмыщелки с бугорками для
связок. Над блоком расположена венечная ямка (рис.
80)
2.7.1.2.
Кости предплечья, запястья, пясти и пальцев
Зейгоподий
представлен костями предплечья: лучевой и локтевой.
Лучевая
кость – os
radii –
относится также к длинным трубчатым костям, расположена впереди от локтевой.
Она имеет слабо выраженную головку с суставной поверхностью, разделенной
гребнем на меньшую латеральную и большую медиальную части. На латеральной
поверхности головки есть лучевая шероховатость для крепления двуглавого
мускула. На проксимальном эпифизе имеются связочные бугорки.
Дистальный
эпифиз несет суставную поверхность для проксимального ряда костей запястья,
разделенную двумя гребнями на три части. С латерального конца выступает
грифельный отросток лучевой кости, а с медиальной – грифельный отросток
локтевой кости.
Локтевая
кость – os
ulna
– срастается с лучевой с латеро-пальмарной поверхности. На проксимальном конце
сильно выражен локтевой отросток с одноименным бугром для крепления трехглавого
мускула, наружная поверхность которого более выпукла. Между головкой луча и
локтевым отростком выражена полулунная вырезка, заканчивающаяся крючковидным
отростком.
На
дистальном конце кость оканчивается грифельным отростком. Между телами лучевой
и локтевой костей имеется межкостное пространство и формируется латеральный
сосудистый желоб (рис. 81).
Кости
запястья (ossa carpi) относятся к базиподию и состоит из двух
рядов костей. В проксимальном ряду у жвачных самая медиальная – лучевая, затем,
промежуточная, локтевая и добавочная (латеральная). В дистальном ряду: вторая +
третья, сросшиеся (медиальная сторона) и четвертая (с латеральной стороны).
Лучевая
кость запястья неправильной кубообразной формы с несколько закругленной
дорсо-медиальной поверхностью. Проксимальная и дистальная поверхности несут
суставные площадки для лучевой кости предплечья и дистального ряда запястья.
Пальмарный край несколько загнут.
Промежуточная
(средняя) кость имеет форму неправильного треугольника с тупыми вершинами.
Дистальная суставная поверхность имеет гребешок, делящий ее на меньшую –
медиальную и большую – латеральную части.
Локтевая
кость запястья крючковидной формы, имеет на проксимальной части седло.
Крючковидный отросток загнут вниз, и несет на себе суставную поверхность для
добавочной кости.
Добавочная
кость удлиненно-округлой формы, имеет суставную поверхность для локтевой
запястной кости и шероховатость для сухожилий.
Вторая
и третья кости запястья срослись в одну кость, лежащую медиально в дистальном
ряду. Кость на проксимальной поверхности разделена гребнем на медиальную
(большую) и латеральную (меньшую) части. Дистальная поверхность плоская и
служит для соединения с пястной костью.
Четвертая
кость запястья лежит латерально рядом с предыдущей, соединяется с локтевой и
промежуточной. На проксимальной поверхности расположен гребешок, делящий ее на
две части. Дистальная поверхность плоская и прилежит к пястной кости (рис. 82).
Кости
пясти (ossa
metacarpi) входят в состав метаподия и состоят из II, III + IV и V костей. На проксимальном конце имеется
суставная поверхность для сочленения с дистальным рядом запястья, которая
разделена гребнем на бóльшую медиальную и меньшую латеральную площадки.
Сбоку проксимального эпифиза расположены связочные шероховатости, а на
дорсальной поверхности имеется пястная шероховатость для лучевого разгибателя
запястья.
Дистальный
эпифиз разделен на два суставных блока для пальцев, которые несут гребни,
разделяющие блоки на большую медиальную и меньшую латеральную площади. Между
блоками расположена глубокая вырезка. С латеральной и медиальной поверхностей
блоков имеются связочные ямки (рис. 83).
Кости
пальцев (ossa digitales). Каждый палец состоит их трех фаланг.
Первая
фаланга (phalanx prima)
относится к трубчатым костям. На проксимальном конце имеется суставная
поверхность, разделенная желобом на две части для блока пястной и плюсневой
костей. Дистальный конец также разделен желобом. Диафиз сверху несет связочные
бугорки (пальмарные и плантарные), а на задней поверхности у дистального
эпифиза расположены связочные шероховатости. Медиальные поверхности фаланг
более уплощены. Первые фаланги этих пальцев грудной конечности короче, чем на
тазовой.
Проксимальный
конец имеет глубокую суставную ямку, дистальный – седловидную суставную
поверхность.
На
дистальном конце расположена суставная поверхность для второй фаланги.
Вторые
фаланги (ph. secunda) на обеих конечностях более
короткие, чем первые. На проксимальном конце имеют суставные поверхности для
первой фаланги, разделенные гребешком, дистальные концы имеют блок с желобом.
На пальмарной и плантарной поверхностях диафизов заметны шероховатости для
связок.
Третьи
фаланги или копытные, копытцевые или когтевые кости (ph. tertia) на опорных пальцах имеют подошвенную
латеральную и медиальную поверхности. Последняя поставлена вертикально
относительно подошвы. На проксимальной части, расположена суставная
поверхность, разделенная гребнем для второй фаланги. Дорсальный край
проксимального конца несет разгибательный отросток, а с пальмарной (плантарной)
стороны заметна фасетка для челночной кости.
Сесамовидные
кости (os sesamoidea)
первой фаланги лежат попарно на пальмарной и плантарной поверхностях пястно- и
плюсно-фаланговых суставов опорных пальцев, имеют суставные поверхности,
разделенные гребнем.
Сесамовидные
кости третьей фаланги (челночные) неправильной четырехугольной формы,
расположены с пальмарной (плантарной) поверхности копытцевого сустава. На
дорсальной суставной поверхности имеются гребни, а дистальная часть несет
фасетки для III
фаланги (рис. 84).
Видовые
особенности:
Лошадь
– на
лопаточной ости находится бугор ости – tuber spina scapulae, лопаточный хрящ
мощный, акромион отсутствует, суставная впадина имеет суставную вырезку.
На проксимальном эпифизе плечевой кости имеется три
бугорка: большой, средниц и малый, межбугоковый желоб двойной. Дельтовидная и
круглая шероховатости, гребено большого бугра хорошо развиты. На блоке имеется
синовиальная ямка.
Лучевая кость хорошо развита, локтевая – редуцирована,
имеется только проксимальный эпифиз, который срастается с лучевой, сохраняется
одно межкостное пространство – spatium
interosseum.
В костях запястья в проксимальном ряду четыре и в
дистальном четыре кости (1-я часто отсутствует).
Имеется три пястные кости, из них хорошо развита 3-я, а
2-я и 4-я рудиментированы и называются грифельными косточками.
Один третий палец. З фаланги: путовая, венечная и копытная.
Крупный
рогатый скот – лопаточная ость заканчивается акромионом,
доходящим до шейки, предостная ямка значительно уже заостной, лопаточный хрящ
большой.
Плечевая кость относительно короткая, большой бугор
вытянут проксимально.
Хорошо развита только лучевая кость, тело локтевой кости
редуцировано. Имеется два межкостных пространства: проксимальное и дистальное.
На дистальном эпифизе локтевой кости имеется шиловидный отросток.
В проксимальном ряду запястья четыре кости, в дистальном
2: срастаются 2 и 3, 4 и 5.
В костях пясти отсутствуют 1 и 2, 3 и 4 срастаются в
одну. На границе их сращения проходят краниальный и пальмарный желоба.
Два пальца. Третьи фаланги – копытцевые.
Свинья –
лопатка короткая и широкая. На лопаточной ости мощный бугор ости, нависающий на
заостную ямку. Акромиона нет, лопаточный хрящ слабо развит.
Плечевая кость короткая и толстая. Большой бугор разделен
надвое. Шероховатости развиты слабо.
Лучевая кость короткая и массивная, тело локтевой кости
также массивное. Обе кости срастаются с помощью волокнистой ткани.
В проксимальном ряду запястья 4 кости, 4 и в дистальном.
4 пястных кости, хорошо развиты.
4 пальца, 3-я – копытцевая.
Собака –
хорошо на лопатке выражен акромион, он достигает суставной впадины. Предостная и
заостная ямки почти равны по ширине. Лопаточный хрящ слабо развит.
Плечевая кость тонкая и длинная, над блоком находится
надблоковое отверстие.
Лучевая и локтевая кости срастаются подвижно.
В запястье в проксимальном ряду 3 кости: срастаются
лучевая и промежуточная, в дистальном 4.
5 или 4 пальца, 3-я фаланга когтевая.
2.7.2.
Скелет тазового пояса и задней конечности
2.7.2.1.
Тазовые и бедренная кости
Тазовый
пояс состоит из двух безымянных костей – os innominata, каждая из которых включает
три кости: подвздошную – os
ilium,
лонную – os pubis и
седалищную – os ishii. В
области суставной впадины все три кости срастаются. Обе безымянные кости
соединяются по сагиттальной линии, в результате формируется тазовый шов.
Важное
значение для прогнозирования родов имеют промеры таза самок. При этом различают
высоту входа в таз (conjugata vera) –
от мыса крестцовой кости до лонного бугра; расстояние между поясничными
бугорками - средний поперечник и расстояние между подвздошно-лонными
возвышениями – вентральный поперечник. Диагональная конъюгата – от мыса
крестцовой кости и до седалищной дуги. Поперечник таза – расстояние между
седалищными остями. Высота выхода из таза измеряется от тела 3-4 хвостового
позвонка и до седалищной дуги в области шва. Ширина выхода – между дорсальными отростками
седалищных бугров.
Все
эти промеры таза у самок на 10-20% больше, чем у самцов, что имеет существенное
значение для развития плода и дальнейших родов. Все бугры и шероховатости
тазовых костей у самок более сглажены.
Подвздошная
кость – os
ilium –
служит основным звеном, соединяющим свободную конечность с крестцовым отделом и
передающим толчки с тазовой конечности на туловище.
Тело
кости направлено дорсо-краниально. Расширенная часть называется крылом, на
котором расположены ягодичные мышцы. Передний край крыла имеет подвздошный
гребень для крепления длиннейшего мускула туловища. Латеральный край крыла
несет наружный подвздошный бугор или маклок, медиальный – внутренний
подвздошный или крестцовый бугор. На латеральной части крыла различают ягодичную
поверхность и ягодичную линию. Медиальная поверхность крыла называется тазовой
и несет ушковидную поверхность для сочленения с крылом крестцовой кости.
На
теле кости с дорсального края различают большую седалищную вырезку, а со
стороны тазовой поверхности – подвздошно-лонный гребень, идущий от ушковидной
шероховатости и до лонной кости. На гребне выделяется поясничный бугорок.
Лонная
кость – os
pubis –
состоит из впадинной и шовной ветвей. Первая участвует в формировании суставной
впадины, вторая направлена в сторону тазового шва. Ветви принимают участие в
образовании запертого отверстия – for. оbturatum.
На
шовной ветви находится подвздошно-лонное возвышение. Шовные ветви правой и
левой костей в области шва образуют лонный бугор.
Седалищная
кость – os
ishii
– состоит из тела и двух ветвей – шовной и впадинной. Так же, как и предыдущая
кость принимает участие в формировании запертого отверстия. Каудально тело
кости имеет седалищный бугор, разделенный на три части, состоящие из
дорсального, латерального и каудального бугорков. Сверху на впадинной ветви
различают малую седалищную вырезку. Между большой и малой седалищными вырезками
расположена седалищная ость для крепления широкой тазовой связки. Между
седалищными буграми образуется седалищная дуга.
Суставная
впадина – acetabulum – служит вместилищем для головки бедренной
кости и образована тазовыми костями. На свежих препаратах края впадины
оформлены хрящевыми губами. На дне впадины имеется ямка для круглой связки, а
на нижнем крае расположена вырезка суставной впадины.
В
целом обе безымянные кости и крестцовая кость образуют тазовую полость, которая
содержит конечные отделы пищеварительного тракта и мочеполового аппарата. У
самок тазовая полость служит, кроме того, вместилищем плода и родовым путем (рис. 85).
Бедренная
кость – os
femoris –
длинная трубчатая кость проксимальным концом образует тазобедренный, дистальным
– коленный суставы.
На
проксимальном эпифизе различают головку, на которой в центре имеется небольшая
ямка для круглой связки. Латерально от головки выступает большой вертел – trochantоr major для закрепления ягодичных
мышц. Медиально расположен малый вертел – trochantоr minor. Между обоими вертелами
имеется межвертлужный гребень – crista
intertrochanterica, ограничивающий вертлужную ямку – fossa trochanterica, где заканчиваются
запирательные мышцы.
На
дистальном эпифизе с дорсальной поверхности имеется блок для коленной чашки – trochlea patellaris, состоящий из медиальной
(большой), латеральной (меньшей) губ и желоба между ними. Заканчивается кость
латеральным и медиальным мыщелками – condylus lateralis et medialis, разделенными межмыщелковой
ямкой.
На
латеральном мыщелке имеется ямка для подколенной мышцы, а между латеральной
губой и мыщелком – разгибательная ямка. На диафизе от большого вертела тянется
слабый гребень (латеральная губа). На дистальной трети диафиза имеется
плантарная ямка или бугорок (рис. 86).
Коленная
чашка – patella –
представлена сесамовидной костью неправильной треугольной формы, которая имеет
суставную поверхность, скользящую по блоку.
На
кости различают основание (верхняя часть) и вершину (нижняя часть).
2.7.2.2.
Кости голени, заплюсны, плюсны и пальцев
Большеберцовая
кость – os
tibia –
относится к длинным трубчатым костям. На проксимальном конце различают
латеральный и медиальный мыщелки, между которыми имеется межмыщелковое
возвышение – eminentia
intercondylaris, состоит из двух бугорков. Медиальный
бугорок чуть больше латерального. Между бугорками проходит межмыщелковый желоб,
разделяя суставную поверхность на медиальную (большую) и латеральную части. В
области желоба расположены две ямки для
крестовидных связок. С плантарной поверхности мыщелки разделены
подколенной вырезкой.
По
дорсальной поверхности на тело кости от мыщелков опускается гребень – crista tibia, достигающий середины
диафиза кости. Между латеральным мыщелком и гребнем выделяется мышечный желоб.
На
дистальном эпифизе имеется блок с суставной поверхностью для таранной кости. Со
стороны медиального блока выступает медиальный лодыжковый отросток – processus malleolus, несущий небольшой желоб
для сухожилия длинного разгибателя пальцев.
С
латеральной стороны блока имеется суставная поверхность для латеральной
лодыжковой кости. Последняя представляет собой неправильной четырехугольной
формы кость для соединения с большеберцовой и костями заплюсны (рис. 27).
Малоберцовая
кость – os
fibula –
как таковая подверглась редукции и представляет тонкую костную пластинку,
расположенную с латеральной поверхности от соответствующего мыщелка и доходящую
до середины tibia.
Дистально она переходит соединительнотканный фиброзный тяж (рис. 87).
Кости
заплюсны состоят из трех рядов костей. В
проксимальном ряду две кости – таранная и пяточная.
Таранная
кость – os
tarsi tibiale (talus) –
относится к коротким костям. Она имеет проксимальный блок для большеберцовой
кости, дистальный для центральной заплюсневой и плантарный для пяточной кости.
Пяточная
кость – os
tarsi fibulare (calcaneus) –
присоединяется с латеральной стороны к предыдущей. Состоит из тела и пяточного
отростка. На теле имеется суставная поверхность для таранной кости, латеральный
и медиальный отростки. Латеральный отросток называется лодыжковым, а медиальный
– держателем таранной кости, на котором выражен желоб для сухожилия глубокого
сгибателя пальцев. На задней поверхности тела пяточной кости имеется плантарный
гребень.
Пяточный
отросток – proc. сalcaneus – направлен проксимально и
заканчивается пяточным бугром, где крепится пяточное сухожилие и проходит
небольшой желоб для поверхностного пальцевого сгибателя.
Средний
ряд костей запястья представлен центральной костью – os tarsi centrale, которая у жвачных
срастается с четвертой и пятой костями и образует центротарсальную кость.
В
дистальном ряду имеется первая, вторая, третья и 4-я+5-я кости заплюсны (рис. 88).
Формула
костей заплюсны выглядит следующим образом:
tt tf___
_t c
t1 t2 t3(t4
+ t5)
Кости
плюсны – ossa
metatarsea – представляют гомодинамные звенья пясти грудной
конечности и состоят из 4 костей или из сросшихся третьей и четвертой плюсневых
костей у жвачных.
На
проксимальном конце имеет плоские суставные поверхности для дистального ряда
костей заплюсны.
Дистальный
эпифиз несет блок, который имеет центральный гребень и два желоба сбоку, а с
плантарной стороны расположены ямки для сесамовидных костей (рис. 89).
Кости пальцев
тазовой конечности сходны по строению с грудными, но имеют некоторые отличия,
которые заключаются в том, что первые и вторые фаланги третьего и четвертого
пальцев массивнее, чем на грудных конечностях. Третьи фаланги несколько короче
передних.
Сесамовидные
кости стопы по внешнему виду сходны с таковыми на передней конечности.
Третьи
фаланги на опорных пальцах имеют подошвенную латеральную и медиальную
поверхности. Последняя поставлена вертикально относительно подошвы. На
проксимальной части, расположена суставная поверхность, разделенная гребнем для
второй фаланги. Дорсальный край проксимального конца несет разгибательный
отросток, а с пальмарной (плантарной) стороны заметна фасетка для челночной
кости.
Сесамовидные
кости (os sesamoidea)
первой фаланги лежат попарно на пальмарной и плантарной поверхностях пястно- и
плюсно-фаланговых суставов опорных пальцев, имеют суставные поверхности,
разделенные гребнем.
Сесамовидные
кости третьей фаланги (челночные) неправильной формы, расположены с пальмарной
(плантарной) поверхности копытцевого сустава. На дорсальной суставной
поверхности имеются гребни, а дистальная часть несет фасетки для III фаланги (рис.
90).
Видовые особенности:
Лошадь
– крылья
подвздошных костей треугольный, налегают на крестец сверху. Крестцовые бугры
сближены, маклок и седалищный бугор пластинчатые с двумя углами, вырезка
суставной впадины глубокая. На дне впадины имеется синовиальная ямка.
На проксимальном эпифизе бедренной кости имеется четыре
вертела: средний, большой, малый и третий. На головке вместо ямки – треугольная
вырезка, медиально расположенная. Плантарная ямка глубокая. Коленная чашка
четырехугольной формы.
На латеральном мыщелке большеберцовой кости имеется
шероховатость для головки малоберцовой кости. На дистальном эпифизе
большеберцовой кости – латеральная и медиальная лодыжки.
Таранная кость заплюсны имеет косо поставленный блок, в
дистальном ряду три кости: 1 и 2 сросшиеся, третья хорошо развита лежит под
центральной, 4-я небольшая.
Кости плюсны и пальцев имеют идентичное строение с
передней коненостью.
Крупный
рогатый скот – седалищная кость высокая, седалищный бугор
несет три бугорка, седалищная дуга глубокая.
Бедренная кость имеет два вертела:большой и малый,
имеется плантарная ямка.
Коленная чашка треугольной формы.
На латеральном мыщелке большеберцовой кости выступает
небольшой бугорок, который является рудиментом головки малоберцовой кости.
Таранная кость имеет два поставленных блока, центральная
кость срастается с 4+5 дистального ряда, в дистальном ряду 3 кости: 1-я, 2+3,
4+5+центральная.
Кости плюсны и пальцев имеют идентичное строение с
передней коненостью.
Свинья –
крыло подвздошной кости узкое и округлое, подвздошный гребень выпуклый маклок и
крестцовый бугор слабо выражены. Большая и малая седалищные вырезки глубокие.
Бедренная кость короткая и массивная, имеет два вертела,
ямка головки маленькая. Имеется плантарная шероховатость.
Большеберцовая кость короткая и массивная. Малоберцовая
кость самостоятельная, развита на всем протяжении, ее дистальный эпифиз имеет
латеральную лодыжку.
Таранная кость несет два блока, в дистальном ряду четыре
кости.
Кости плюсны и пальцев имеют идентичное строение с
передней коненостью.
Собака –
крыло подвздошной кости ложечковидное, подвздошный гребень дугообразный, маклок
и крестцовый бугор не выражены. Малая седалищная вырезка плоская.
Бедренная кость тонкая и длинная. Большой вертел
выступает над головкой. Имеется плантарный бугорок.
Большеберцовая кость длинная и тонкая. Малоберцовая
развита на всем протяжении.
Кости плюсны и пальцев имеют идентичное строение с
передней коненостью.
2.8.
Соединение
костей
СИНДЕСМОЛОГИЯ sindesmologia –
учение о соединении костей. Соединения
костей объединяют кости скелета в единое целое. Они удерживают их друг возле
друга и обеспечивают им большую или меньшую подвижность. Соединения костей
имеют различное строение и обладают физическими свойствами, как прочность,
упругость, подвижность, что связано с выполняемой ими функцией.
2.8.1.
Виды
соединения костей
1. Непрерывные
соединения, в которых между костями имеется
прослойка соединительной ткани или хряща. Щель или полость между соединяющимися
костями отсутствует.
2. Прерывные соединения,
или суставы (синовиальные соединения),
характеризуются наличием между костями полости и синовиальной мембраны,
выстилающей изнутри суставную капсулу.
3. Симфизы, или
полусуставы, имеют небольшую щель в хрящевой или
соединительнотканной прослойке между соединяющимися костями (переходная форма
от непрерывных соединений к прерывным).
2.8.2.
Непрерывные
соединения
Синартроз – это соединение 2-х
костей, пространство между которыми заполнено тканью, выполняющей соединяющую
роль.
1. Синдесмоз – это соединение костей с помощью соединительной
ткани (Эта ткань характеризуется наличием большого количества коллагеновых
волокон) (рис. 91).
Разновидности:
связка – лентовидное подвижное соединение.
мембрана – пластинчатое подвижное соединение (например,
соединение между атлантом и затылочной костью)
швы – неподвижные соединения плоских костей: плоский,
зубчатый, листочковый, чешуйчатый, вколачивание (соединение зубов с костью).
2. Синсаркоз – это соединение костей при помощи мышечной ткани
(например, соединение рёбер между собой) (рис. 92).
3. Синэластоз – соединение костей при помощи соединительной ткани,
в которой преобладают эластические волокна (рис. 93).
4. Синхондроз – соединение костей при помощи хрящевой ткани (у
молодых животных так соединяются грудная кость, эпифиз с диафизом,
тазовые кости, кости черепа) (рис. 94).
5. Синостоз – соединение костей при помощи костной ткани (у
взрослых животных является сменной стадией развития скелета на месте
соединительной и хрящевой ткани формируется костная ткань, например, крестцовая
кость) (рис. 95).
2.8.3.
Прерывные
соединения (суставы, диартрозы)
Диартроз – это
такой вид соединения кости, при котором между костями имеется полость,
заполненная синовиальной жидкостью (рис. 96).
Основные компоненты сустава:
1. Гиалиновый хрящ – cartilage articularis, покрывающий суставную поверхность соединяющихся
костей Синовиальная жидкость
2. Суставная капсула. Состоит из 2-х слитков: наружный
слой называется фиброзный, выполняет защитную функцию, состоит из
соединительной ткани, внутренний – синовиальный, состоит из клеток,
расположенных в один слой и вырабатывающих синовиальную жидкость
3. Суставная полость
4. Костная ткань
5. Классификация
суставов:
1. по строению простые и сложные. простой – между
соединяемыми поверхностями нет включений, сложных – между соединяемыми
поверхностями есть включения (диски, мениски, короткие кости) (рис. 97)
2. по осям вращения: одноосные (локтевой, коленный,
суставы пальцев), двухосные (затылочно-атлантный, височно-челюстной),
многоосные (плечевой, тазобедренный)
3. по форме соединяемых поверхностей: блоковидные,
цилиндрические, эллипсовидные, шаровидные, плоские (рис.
98)
Движение в суставах:
Сгибание
- flexio,
Разгибание
- exstensio,
Вращение
- rotatio, вращение вовнутрь - pronatio, вращение наружу - supinatio, круговое движение - circumductio
отведение
- abductio,
приведение
- adductio.
Имеются
тугие суставы, в которых движение практически отсутствует
(крестцово-подвздошный сустав)
2.8.4.
Соединения
осевого скелета
Соединение
костей черепа.
1.
швы (рис. 99)
2. синхондроз (затылочная кость с клиновидной)
3. синостоз (у взрослых животных)
4. вколачивание (зубы в альвеолах)
5. простые суставы (соединения члеников подъязычной кости
между собой)
Височно-челюстной
сустав образован суставным бугорком височной кости и суставными отростками
нижнечелюстной кости. Сустав двуосный (к.р.с., лошадь) одноосный (свинья,
собака), сложный, эллипсовидный. В суставе имеется капсула, боковые связки,
каудальная связка (к.р.с., лошадь) (рис. 100)
Соединения
позвонков:
Атлантно-затылочный
сустав. Образован мыщелками затылочной кости, и краниальными суставными
поверхностями атланта. сустав простой, 2-хостный, эллипсовидный. 2 капсулы, 2
мембраны и 2 боковые связки.
Ось-атлантный сустав.
Образован зубовидной ямкой атланта, каудальными суставными поверхностями
атланта, а также зубовидным отростком и краниальными суставными поверхностями
эпистрофея. сустав простой, цилиндрический, одноосный. В суставе имеются
капсулы, дорсальная мембрана, дорсальная связка зубовидного отростка,
поперечная связка (собака, свинья), вентральная связка зубовидного отростка
(к.р.с., лошадь) (рис. 101)
Позвонки
отделов со 2-го шейного и крестцовая кость имеют следующие соединения:
1. соединения тел позвонков: межпозвоночные диски
(синхондроз) (рис. 102), дорсальная продольная
связка (от 1-го шейного до крестцовой кости), вентральная продольная связка (от
последнего грудного до крестцовой кости).
2. дужки позвонков соединяются синэластозом
3. остистые отростки соединяются синэластозом
(надостистые и межостистые связки)
4. краниальные и каудальные суставные поверхности
простыми плоскими суставами.
Надостистая
связка в краниальном направлении имеет пластинчатую и канатиковую части
Видовые
особенности выйной связки:
У к.р.с.
имеется канатиковая и пластинчатая части, у лошади пластинчатая часть
раздваивается, у свиньи выйная связка отсутствует, у собаки есть только
канатиковая часть. На грудных позвонках имеются соединения позвонков с рёбрами.
Поясничные
позвонки. Поперечные отростки соединяются между собой межпоперечными связками.
У лошади между поперечными отростками 5 и 6 поясничных позвонков и крыльями
крестцовой кости имеются межпоперечные суставы.
Крестцовые
позвонки соединяются у молодых животных – синхондрозом, у старых – синостозом.
Крестцовая кость, соединяясь с подвздошной костью формируют
крестцово-подвздошный сустав, обеспечивая присоединения тазовой конечности к
туловищу. Сустав простой, плоский, тугой, имеется капсула и 4 связки.
Суставная головка ребра и сустав бугорка ребра. Здесь имеются капсулы и связки:
1. Радиальная связка головки
2. Связка шейки ребра
3. Межреберная связка первого сегмента
Соединения рёбер.
1. Рёбра дорсально соединяются с грудными позвонками
2. Рёбра соединяются между собой синсаркозом и
синдесмозом
3. костное ребро соединяется с рёберным хрящом
синартрозом и диартрозом со 2 по 1 у к.р.с. и с 6 по 9 у свиньи.
4. рёберные хрящи истинных рёбер с грудной костью
соединяются грудинно-рёберными суставами. Имеется капсула и ростральная связка.
5. Рёберные хрящи ложных рёбер соединяются между собой
синдесмозом (рис. 103).
Соединения грудины: грудная кость соединяется с грудными хрящами ложных
рёбер; сегменты тела грудины соединяются между собой синхондрозом (у молодых) и
синостозом (у взрослых); у к.р.с. и у свиней рукоятка грудной кости с телом
соединяется суставом, и здесь имеются связки грудины.
2.8.5.
Соединение
костей периферического скелета
2.8.5.1.
Грудная
конечность
Грудная конечность прикрепляется к
туловищу синсаркозом.
n Соединение лопатки с плечевой костью образует плечевой сустав: простой, многоостный,
шаровидный, имеется капсула сустава (рис. 104).
Движения:
разгибание, сгибание, отведение, приведение, вращение
n Соединение плечевой кости: соединения диафиза и эпифиза – синхондроз у молодых,
синостоз у взрослых.
n Локтевой сустав (образован блоком плечевой кости, и проксимальным
эпифизом костей предплечья. Сустав простой (лошадь, свинья, к.р.с) сложный у
собаки; одноостный, блоковидный. Имеются капсула и связки (боковые латеральная
и медиальная). У собак кроме них –
кольцевая (удерживает локтевую кость около лучевой), локтевая (соединяет
медиальный край локтевого отростка с надмыщелком плечевой кости). У лошади -
длинная медиальная связка; Движения:
разгибание, сгибание (рис. 105)
n Соединения костей
предплечья: Диафизы с эпифизом синхондроз синостоз.
Локтевая с лучевой. Синхондроз, синостоз (лошадь, к.р.с.) синдесмоз (у свиньи)
диартроз ( у собаки). Связки: межкостная предплечья, поперечные латеральная и
медиальная.
n Запястный сустав. Сустав сложный, одноостный, блоковидный. образован
костями предплечья, 2-мя рядами костей запястья и костями пястей, в свою
очередь состоит из 3-х суставов (предплечно-запястный, межзапястный, и
запястно-пястный. Связки: боковые длинная и короткая медиальные и латеральные,
межкостные, межрядовые и связки добавочной костиДвижения: разгибание, сгибание (рис. 106)
n Соединения костей пясти. Пястные кости соединены между собой синостозом у
к.р.с. синдесмозом у лошади и свиньи, синсаркозом (у собаки).
2.8.5.2. Тазовая конечность
n
Тазовые кости
обеспечивают соединение тазовой конечности с туловищем с помощью
крестцово-подвздошного сустава – простой, тугой и образуется ушковидными
поверхностями крестца и подвздошной кости. Имеются капсула и связки:
подвздошно-крестцовая вентральная, дорсальная (короткая), межкостная (длинная),
крестцово-седалищная (рис. 107).
n
Кости
подвздошная, седалищная и лонная срослись между собой синхондрозом и синостозом
и формирует тазовую кость
n
Тазобедренный
сустав образован суставной впадиной тазовой кости и головкой бедренной кости.
Сустав простой, многоосный, чашеобразный. Связки: капсула сустава, круглая,
поперечная связка ацетабулюм и добавочная (у лошади – направляется от лонного
бугра к ямке головки, она препятствует отведению конечности в бок).
n
Движения:
разгибание, сгибание, отведение, приведение, и вращение – супинация (наружу) и
пронация (внутрь).
n Коленный сустав образован дистальным эпифизом бедренной кости и
проксимальным эпифизом костей голени. сустав сложный, одноостный, блоковидный.
Сложный из за наличия менисков. коленный сустав состоит из 2-х суставов: а)
бедренно-чашечный (сустав коленной чашечки) б) бедренно-большеберцовый. Связки:
капсула сустава, боковые латеральная и медиальная, крестовидные, связки
менисков и коленной чашки.Движения: разгибание,
сгибание (рис. 108)
n Заплюсневый сустав образуется костями голени, заплюсны и плюсны. Связки:
капсула, боковые длинные и кроткие, межрядовые, длинная (заплюсневая)
плантарная и дорсальная заплюсневая, межкостныеДвижения: разгибание, сгибание
а) бедренно-чашечный (сустав коленной
чашечки) б) бедренно-большеберцовый (рис. 109).
Суставы пальцев: путовый,
венечный, копытный – простые, одноосные. Связки: капсула сустава, боковые
медиальная и латеральная, связки сесамовидных костей (рис.
110)
2.9.
Скелетная
мышечная система у животных
Миология (Myologiа) - это раздел анатомии животных, изучающий
строение мышечной системы
2.9.1.
Общие
сведения о скелетной мышечной системе животных
Мышечная
ткань, составляющая основу этой системы, осуществляет все двигательные процессы
в организме животных. Благодаря ей тело фиксируется в определенном положении и
перемещается в пространстве, осуществляются дыхательные движения грудной клетки
и диафрагмы, движение глаз, глотание, двигательные функции внутренних органов,
включая работу сердца (рис. 111).
Почему
кошка всегда падает на четыре лапы?
Долгое время полагали, что вращающееся вокруг оси и
ни на что не опирающееся тело не может повернуться в пространстве одними лишь
внутренними усилиями. Загадка кошачьего падения была решена в связи с вопросом
об искусственном перемещении полюсов. У кошки есть два средства повернуть свое
тело при падении. Первое средство, это – перемещение хвоста: когда кошка, держа
хвост под углом к своему телу, производит им вращательное движение, то все тело
немного поворачивается в обратном направлении. Рядом последовательных оборотов
хвоста кошка может повернуть свое тело на желаемый угол. Во-вторых, кошка при
повороте вытягивает и укорачивает передние и задние лапы: согласно так
называемому закону площадей часть тела с вытянутыми лапами должна, при равных
прочих условиях, повернуться на меньший угол, нежели часть тела с прижатыми
лапами. Чередуя надлежащим образом вытягивание и прижатие лап, кошка может
рядом телодвижений достичь нужного поворота в желаемом направлении (рис. 112)
У саранчи мышц до 900,
а у некоторых гусениц — до 4 тыс.
Самым быстрым сухопутным
животным является гепард. На дистанции до 500 м он может развивать скорость до
101 км/ч. Лучшая скаковая лошадь движется со скоростью 72 км/ч.
Самая быстрая рыба —
тихоокеанский парусник — на дистанции в 100 м сумела развить скорость до 109
км/ч.
Что же касается силачей в природе, то муравей, например,
может поднимать челюстями вес, в 50 раз больший, чем вес самого насекомого. А
жук-носорог умудряется удержать на спине вес, в 850 раз больший, чем его
собственный. Для сравнения: слон может удержать на спине только четверть своего
веса. Общая закономерность такова: чем больше животное, тем меньший вес в
сравнении с собственным оно может поднять
Мышечная
ткань обладает специальными сократительными органеллами - миофибриллами.
Миофибриллы, состоящие из тонких белковых нитей (миофиламентов), могут быть
неисчерченными или исчерченными (поперечнополосатыми). Соответственно различают
неисчерченную и исчерченную мышечную ткань (рис. 113).
Неисчерченная мышечная ткань состоит из клеток (гладких миоцитов) веретеновидной
формы. Эти клетки образуют мышечные слои в стенках кровеносных и лимфатических
сосудов, в стенках внутренних органов (желудок, кишечник, мочевыводящие пути,
матка и т.д.). Длина клеток колеблется от 20 мкм (в стенке кровеносного сосуда)
до 500 мкм (в стенке матки стельной коровы), диаметр от 2 до 20 мкм. В
функциональном отношении неисчерченная мышечная ткань имеет ряд особенностей:
она обладает большой силой (например, в кишечнике постоянно передвигаются
значительные массы пищи), обладает слабой утомляемостью, медленным сокращением
и ритмичностью движений (в стенке кишечника неисчерченная мышечная ткань
сокращается 12 раз в минуту, а в селезенке - только 1 раз). Сокращение гладких
мышц происходит непроизвольно (рис. 114).
Исчерченная
мышечная ткань характеризуется наличием исчерченных миофибрилл, имеет 2
разновидности.
А) Исчерченная сердечная мышечная ткань
состоит из удлиненных клеток (кардиомиоцитов) квадратной формы. Их концы, соединяясь
друг с другом в цепочки, формируют так называемые функциональные мышечные
"волокна" толщиной 10-20 мкм. Тесно связываясь между собой,
функциональные мышечные "волокна" образуют мышечную оболочку сердца
(миокард), постоянные и ритмичные сокращения которого приводят в движение кровь
(рис. 115).
Б) Исчерченная скелетная
мышечная ткань, в отличие от сердечной, состоит не из клеток, а из многоядерных
мышечных образований (миосимпластов) цилиндрической формы. Длина миосимпластов
колеблется от нескольких миллиметров до 13-15 см, диаметр от 10 до 150 мкм.
Количество ядер в них может достигать нескольких десятков тысяч. Миосимпласты
(их еще называют "мышечными волокнами") образуют скелетные мышцы и
входят в состав некоторых органов (язык, глотка, гортань, пищевод и др.). В
функциональном отношении скелетная мышечная ткань легко возбудима и сокращается
быстрее, чем неисчерченная (например, в обычных условиях скелетная мышца
сокращается в течение 0,1 с, а неисчерченная - в течение нескольких секунд).
Но, в отличие от гладких (неисчерченных) мышц внутренних органов, скелетные
мышцы быстрее утомляются (рис. 116).
СТРОЕНИЕ МЫШЦЫ КАК ОРГАНА
Скелетная мышца (Musculus
skeleti) - это активный орган аппарата движения, форма и особенности строения
которого обусловлены выполняемой функцией и местоположением на скелете. В мышце
различают активно сокращающуюся часть - мышечное брюшко и пассивную часть, при
помощи которой она прикрепляется к костям, - сухожилие.
В мышце
выделяют головку (caput) — начальную часть, брюшко (venter) — среднюю часть и
хвост (cauda) — конечную часть. От длины мышцы зависит степень размаха, который
она может обеспечить. У каждой мышцы есть точка начала (origo) и место
крепления (insertio). В мышце
выделяют головку (caput) — начальную часть, брюшко (venter) — среднюю часть и
хвост (cauda) — конечную часть. От длины мышцы зависит степень размаха, который
она может обеспечить. У каждой мышцы есть точка начала (origo) и место
крепления (insertio) (рис. 117).
1)
Мышечное брюшко (venter) состоит из паренхимы и стромы.
Паренхима представлена исчерченной мышечной тканью, структурной единицей
которой является миосимпласт. Миосимпласты объединяются при помощи рыхлой
соединительной ткани, которая называется эндомизий, в пучки 1 порядка. Пучки 1
порядка объединяются в пучки 1,2,3 порядка и между ними формируются
соединительнотканные перегородки (перимизий), по которым внутрь в мышцу
проникают сосуды и нервы. Снаружи мышечное брюшко покрыто соединительнотканной
оболочкой (эпимизием). Эндо-, пери- и эпимизий образуют строму мышечного брюшка
и защищают мышцу от чрезмерного утолщения или растяжения. Соединительнотканные
элементы, имеющиеся между мышечными волокнами, по концам мышечного брюшка,
переходят в сухожилия.
2)
Сухожилие (tendo) построено
по тому же принципу, что и мышечное брюшко, с той лишь разницей, что вместо
мышечных волокон его пучки содержат коллагеновые волокна. Прослойки
соединительной ткани внутри носят названия эндо- и пери теноний, а снаружи
плотная соединительная ткань образует оболочку (эпитеноний), которая является
продолжением эпимизия. Сухожилие имеет блестящий светло-золотистый цвет, резко
отличающийся от красно-бурого цвета брюшка мышцы. В большинстве случаев сухожилие
находится по обоим концам мышцы и прикрепляется к костям. Хотя сухожилие
значительно тоньше мышечного брюшка, прочность его велика, оно способно
выдерживать большую нагрузку и практически нерастяжимо. Исследования показали,
что для разрыва ахиллового сухожилия у животного требуется сила от 900 кг на
один кубический см (рис. 118).
3)
Сосуды и нервы входят в мышцу с
ее внутренней стороны.
- Артерии ветвятся до капилляров,
которые в пучках мышечных волокон образуют густую сеть. К каждому мышечному
волокну прилежит не менее одного кровеносного капилляра. В каждую мышцу кровь
поступает по артериям, а оттекает по венам и лимфатическим сосудам.
- Нервы, разветвляясь в мышце, образуют
нервно-мышечный комплекс – мион, который состоит из 1 нервного волокна и
нескольких мышечных волокон. Так, например, в трехглавой мышце голени мион
состоит из 1нервного волокна и 227 мышечных волокон, а в латеральной мышце
глаза – из 1 нервного волокна и 19 мышечных волокон (рис.
119).
Рост мышц в длину происходит в так называемых «зонах
роста», которые располагаются в местах перехода мышечного брюшка в сухожилие и
содержат большое количество ядер, а увеличение мышц в толщину происходит
благодаря функциональной нагрузке, которую выполняет данная мышца (рис. 120).
2.9.2.
Особенности
исторического и индивидуального развития скелетных мышц
Большинство
многоклеточных животных движется с помощью мышечных сокращений. «Смеётся ли
ребёнок при виде игрушки, улыбается ли Гарибальди, когда его гонят за излишнюю
любовь к родине, создаёт ли Ньютон мировые законы и пишет их на бумаге — везде
окончательным фактом является мышечное движение» (Иван Сеченов). Этот список
можно продолжить, добавив сюда почти любые движения большинства многоклеточных
животных. Насколько разнообразны эти движения, основанные на общем принципе!
Учёные выделяют
несколько типов мышц. Наиболее древняя по происхождению мышечная ткань
называется гладкой. У большинства беспозвоночных (например, моллюсков)
имеется только гладкая мускулатура. Сокращаются такие мышцы очень медленно (не
случайно улитки стали «образцом» неторопливости). Зато и устают эти мышцы тоже
медленно. Мышца, сжимающая створки раковины моллюска, к примеру, может
оставаться в состоянии сокращения много дней подряд.
У человека гладкие
мышцы, в частности, сужают кровеносные сосуды, создают волнообразные движения
кишечника, регулируют ширину зрачка, поднимают волосы и образуют «гусиную
кожу». Сознательно управлять этими движениями человек не может.
У членистоногих и
позвоночных в ходе эволюции независимо друг от друга возник новый тип мышц. Их
называют скелетными, или поперечнополосатыми. Профессор Николай Бернштейн писал
по этому поводу: «Условия борьбы за существование, конкуренция между живыми
тварями постепенно становились всё жёстче и злее. Жизнь уже не могла мириться с
медлительными, мягкотелыми организмами, рыхлыми, как студень, и подвижными,
вроде часовой стрелки. Борьба и отбор требовали новых исканий.
Поперечнополосатая
мышца полностью решила проблему быстроты и мощности — того, чего так жестоко не
хватало древним мягкотелым. Мышца нового типа способна сокращаться с
молниеносной быстротой: вспомним хотя бы движения крыльев мухи или комара,
совершаемые с частотою нескольких сотен в секунду. При этом, сокращаясь, мышца
легко развивает высокую мощность, в тысячи раз превосходящую древние мышечные
клетки (гладкие мышцы)».
Действительно,
скелетная мышца могла сокращаться порой в тысячу раз быстрее, чем гладкая. Но
зато скелетные мышцы так же быстро и уставали.
В клетках простейших,
губок и кишечнополостных встречаются сократительные волокна, но
специализированные мышечные клетки появляются лишь у плоских и круглых червей.
У всех беспозвоночных до уровня моллюсков они лишены поперечной исчерченности и
напоминают клетки гладкой мускулатуры позвоночных. Сокращаются они не очень
сильно и всегда относительно медленно. Исключение здесь составляют моллюски:
мышцы-замыкатели у двустворок могут рассматриваться как скелетные (рис. 121).
Развитая мускулатура присуща
кольчатым червям, особенно земляным. В стенке их тела находятся кольцевые
мышцы, которые уменьшают его диаметр, и продольные, его
укорачивающие. Здесь же находятся микроскопические мышцы (в каждом
сегменте тела их 4 пары), двигающие щетинками и способные вонзать их в почву.
Земляной червь ползет характерным для него способом за счет сокращений всех
трех категорий мышц – кольцевых, продольных и микроскопических (рис. 122).
Превосходная поперечнополосатая
мускулатура, способная к быстрому и мощному сокращению, характерна для членистоногих.
Летательные мышцы у некоторых насекомых – наиболее быстродействующие из
всех известных: они превосходят в этом смысле даже аналогичные мышцы колибри.
Интересно отметить, что скелетная мускулатура членистоногих управляет движениями
наружного скелета, находясь внутри него, под его защитой (рис. 123).
У ланцетника она
представлена парной продольной мышцей (правой и левой), которая идет
вдоль тела и разделяется соединительнотканными перегородками (миосептами)
на короткие прямые мышечные пучки (миомеры). Такое (сегментарное)
деление единого мышечного пласта называется метамерией (рис.
124)
У круглоротых и рыб
сегментарные мышцы остаются в своем исходном и наиболее
элементарном состоянии. В плавниках рыб они устроены просто и состоят в
основном из поднимателей и опускателей. В конечностях четвероногих они
многочисленны и разнообразны по функциям. Сегментарные мышцы прикрепляются к
костям скелета либо непосредственно, либо при помощи сухожилий (тяжей
соединительной ткани) (рис. 125).
С выходом на сушу и
увеличением разнообразия движений у амфибий и рептилий происходит
разделение дорсальной мышцы, так же как и вентральной, на два тяжа: латеральный
(поперечно-реберная мышца) и медиальный (поперечно-остистая м.). Кроме
этого, у рептилий из латерального тяжа впервые появляются подкожные мышцы,
которые прикрепляются к коже (рис. 126).
У более высокоорганизованных
животных (птиц и млекопитающих) происходит дальнейшая дифференциация
мышечной системы: латеральный и медиальный тяжи, каждый из них, разделяются
на два слоя (поверхностный и глубокий). Кроме этого, у млекопитающих впервые
появляется диафрагма (рис. 127).
В
онтогенезе мышечная система в основном развивается из миотомов мезодермы,
исключение составляют некоторые мышцы головы и шеи, которые образуются из
мезенхимы (трапециевидная, плечеголовная).
В начале формируется мышечный
продольный тяж, который сразу же дифференцируется на дорсальный и вентральный
пласты; далее каждый из них разделяется на латеральный и медиальный пласты,
которые, в свою очередь, дифференцируются на поверхностный и глубокий слои,
последние дают начало определенным группам мышц. Например, из поверхностного
слоя латерального пласта развиваются подвздошно-реберные мышцы, а из глубокого
слоя латерального пласта – длиннейшие мышцы спины, шеи, головы (рис. 128).
2.9.3.
Классификация
мышц
Мышечную систему в
зависимости от особенностей строения, характера двигательной функции и
иннервации делят на соматическую и висцеральную.
Соматическая мышечная
система составляет 40% от массы тела и построена из миосимпластов. Она
произвольная и иннервируется соматической нервной системой. Соматические мышцы
сокращаются быстро, энергично, но кратковременно и быстро утомляются. Такой тип
сокращения называется тетаническим и он характерен для соматической
мускулатуры. К ней относятся:
1) подкожные мышцы, которые не имеют
связи со скелетом и прикрепляются к коже; их сокращения вызывают подергивания
кожи и позволяют ей собираться в мелкие складки;
2) скелетная мускулатура, которая
закрепляется на скелете;
3) диафрагма – куполообразная мышца,
отделяющая грудную полость от брюшной;
4) мышцы языка, глотки, гортани, ушной
раковины, глазного яблока, среднего уха, пищевода и наружных органов
размножения.
Висцеральная мышечная
система составляет 8% от массы тела и построена из гладких миоцитов. Она
непроизвольная и иннервируется вегетативной нервной системой. Гладкие мышцы
сокращаются медленно, длительно и не требуют большого количества энергии. Такой
тип сокращения называют тоническим и он характерен для висцеральной
мускулатуры, которая образует мышечные пучки, слои и оболочки внутренних органов.
Классификация
мышц:
По
форме:
Длинные
мышцы соответствуют длинным рычагам движения и поэтому встречаются главным
образом на конечностях. Имеют веретенообразную форму, средняя часть называется
брюшком, конец, соответствующий началу мышцы, - головкой, противоположный конец
- хвостом. Сухожилие длинных мышц имеет форму ленты. Некоторые длинные мышцы
начинаются несколькими головками (многоглавые) на различных костях, что
усиливает их опору. Встречаются мышцы двуглавые (двуглавая м. плеча),
трехглавые (трехглавая м. голени) и четырехглавые (четырехглавая м бедра). По
количеству брюшек – двубрюшная.
2) Короткие мышцы находятся на тех участках
тела, где размах движений невелик (между отдельными позвонками (многораздельные
м.), между позвонками и ребрами (подниматели ребер) и т.д.).
3) Плоские (широкие) мышцы располагаются
преимущественно на туловище и поясах конечностей. Они имеют расширенное
сухожилие, называемое апоневрозом. Плоские мышцы обладают не только
двигательной функцией, но также опорной и защитной (например, мышцы брюшной
стенки защищают и способствуют удержанию внутренних органов).
4) Встречаются также и другие формы мышц:
квадратная, круговая, дельтовидная, зубчатая, трапецевидная, веретеновидная и
др.
(рис. 129)
По анатомическому строению мышцы делятся в зависимости от количества
внутримышечных сухожильных прослоек и направления мышечных прослоек:
— Одноперистые. Для них характерно
отсутствие сухожильных прослоек и мышечные волокна присоединяются к сухожилию
одной стороны (наружная косая брюшная м.).
— Двуперистые.
Для них характерно наличие одной сухожильной прослойки и мышечные
волокна присоединяются к сухожилию с двух сторон (трапецевидная м.).
— Многоперистые.
Для них характерно наличие двух и более сухожильных прослоек, в
результате этого мышечные пучки сложно переплетаются и к сухожилию подходят с
нескольких сторон (жевательная м., дельтовидная мышца) (рис.
130)
Физиологические и
морфологические поперечники мышц
— Анатомический поперечник –
это площадь поперечного сечения мускула в наиболее широком месте. разрез идёт
перпендикулярно длинной оси мускула
— Физиологический поперечник – это
площадь поперечного сечения мускула, но разрез проводится перпендикулярно
мышечному волокну (рис. 131)
По гистоструктуре все мышцы делятся на 3 типа в зависимости от
соотношения исчерченной мышечной ткани к соединительной
1)
Динамический тип. Для динамических мышц, обеспечивающих активную и
разностороннюю работу, характерно значительное преобладание исчерченной
мышечной ткани над соединительной (четырехглавая м. бедра).
2)
Статический тип. В отличие от динамических, статические мыщцы совсем не имеют
мышечных волокон. Они выполняют большую статическую работу при стоянии и опоре
конечности о почву во время движения, закрепляя суставы в определенном
положении (третья межкостная м. коровы и лошади). Физиологический поперечник больше
анатомического.
3) Статодинамический тип. Для этого типа
характерно уменьшение отношения исчерченной мышечной ткани к
соединительно-тканным элементам (двуглавая м. плеча лошади). Статодинамические
мышцы, как правило, имеют перистое строение. Физиологический поперечник больше
анатомического.
По функции:
1. Флексоры, или сгибатели, которые при сокращении сближают концы
костей
2. Экстензоры, или разгибатели, которые проходят через вершину угла
сустава и при сокращении раскрывают его.
3. Абдукторы,
или отводящие мышцы, лежат на латеральной стороне сустава и отводят его от
сагиттальной плоскости в сторону.
4. Аддукторы,
или приводящие мышцы, лежат на медиальной поверхности сустава и при сокращении
приводят его к сагиттальной плоскости.
5. Ротаторы,
или вращатели, обеспечивающие вращение конечности наружу (супинаторы) или во внутрь (пронаторы).
6. Сфинктеры,
или запиратели, которые располагаются вокруг естественных отверстий и при
сокращении закрывают их. Для них, как правило, характерно круговое направление
мышечных волокон (например, круговая мышца рта).
7. Констрикторы,
или суживатели, которые также относятся к типу круглых мышц, но имеют иную
форму (например, констрикторы глотки, гортани).
8. Дилататоры,
или расширители, при сокращении открывают естественные отверстия.
9. Леваторы,
или подниматели при сокращении поднимают, например ребра.
10. Депрессоры,
или опускатели.
11. Тензоры,
или напрягатели, своей работой напрягают фасции, не позволяя им собираться в
складки.
12. Фиксаторы,
укрепляют сустав на стороне расположения соответствующих мышц.
Мышцы действуют раздельно
или группой. Одинаково действующие мышцы называются синергистами, а действующие противоположным образом - антагонистами.
По отношению к суставам:
1. Односуставные действуют только на один
сустав (предостная м., заостная м. действуют на плечевой сустав).
2. Двусуставные действуют на два сустава
(напрягатель широкой фасции бедра действует на тазобедренный и коленный
суставы).
3. Многосуставные (двуглавая м. бедра, полусухожильная
м., полуперепончатая м. действуют на 3 сустава (тазобедренный, коленный,
скакательный).
2.9.4.
Механизм
мышечного сокращения
Состоят мышцы из
мышечных волокон. Благодаря чему они сокращаются? Долгое время ответ на этот
вопрос оставался для учёных загадкой. Мы знаем, что из всех веществ организма
белки наиболее способны к разным удивительным превращениям. Быть может, при
сокращении мышц уменьшается длина белковых молекул?
Учёным было известно,
что в состав мышечных волокон входят нити двух видов: тонкие (из белка актина)
и толстые (из белка миозина). В 1954 г. ряд биологов одновременно пришли к
выводу, что толстые и тонкие нити не меняют своей длины, но могут скользить
друг относительно друга.
В скелетной мышце
толстые и тонкие нити очень правильно чередуются. Подобно зубьям двух
гребёнок, они входят друг в друга. Под электронным микроскопом те участки, где
«гребёнки» входят друг в друга, выглядят тёмными, а где они разъединены —
светлыми. Получается правильное чередование тёмных и светлых полос. За это
скелетные мышцы и назвали поперечнополосатыми.
Что же касается гладких
мышц, то в них тоже есть тонкие и толстые нити актина и миозина, но они довольно
беспорядочно перемешаны. Поэтому и чередования полос под микроскопом нельзя увидеть.
Иннервация
мускула осуществляется нервами, которые в своём составе содержат следующие виды
нервных волокон:
1.
чувствительные (от мускула к стенке кровеносного сосуда)
2.
двигательные (соматическая нервная система, обеспечивает контроль над работой
мышечного волокна)
3.
двигательные (симпатическая нервная система, обеспечивает иннервацию стенки
сосудов)
2.9.5.
Функции
мышц
Способы движения
четвероногих. Самый простой способ движения — медленный шаг. При шаге в каждый
момент времени приподнята только одна нога, остальные три служат опорой. При
более быстрых аллюрах (способах хода и бега) опора на три ноги исчезает, в
каждый момент движения тело опирается не более чем на две ноги.
Многие хищники и
копытные свободно выбирают наилучший аллюр для каждой скорости. Для лошади,
например, при скорости движения до 5 км/ч наилучший аллюр — шаг. При скорости 7
км/ч выгоднее перейти на рысь, а начиная с 15 км/ч, самым удобным аллюром
становится галоп. Правда, спортивным рысакам приходится часто и при скорости 45
км/ч передвигаться рысью.
Мышца — более
эффективный двигатель, чем, к примеру, двигатель внутреннего сгорания, не
говоря уж о паровом. Её КПД достигает 50% (у автомобиля — не выше 35%).
Кстати говоря, и остальная энергия не пропадает, поддерживая температуру тела.
Известно, что лучший способ согреться — поработать мышцами. Когда человеку
холодно, мышцы начинают непроизвольно сокращаться (тогда человек дрожит),
выделяя тепло.
Чтобы мышца работала,
необходимо «топливо». Об универсальном клеточном топливе (АТФ) рассказано в
статье «Вещества организма». Если его запас иссякает, у человека возникает
чувство изнеможения, «слабости в коленках», когда кажется, что мышцы отказались
служить.
Энергия нужна не только
для сокращения, но и для расслабления мышц. Если запас «топлива» исчерпан,
например, у погибших животных, мышцы «окоченевают», не способные ни
расслабиться, ни сократиться.
Общие закономерности
расположения мышц на конечности
1. Обычно мышцы всегда
лежат выше того сустава, на который они действуют.
2. Мышцы-флексоры,
сгибающие сустав, всегда располагаются внутри сустава, а мышцы-экстензоры
всегда проходят через вершину угла сустава.
3. Отводящие
мышцы-абдукторы всегда лежат на латеральной поверхности, а приводящие
мышцы-аддукторы – на медиальной.
4. Мышцы-супинаторы, вращающие
сустав наружу, лежат ближе к латеральной поверхности, а мышцы-пронаторы,
вращающие сустав во внутрь,- ближе к медиальной поверхности.
5. На костях зейгоподия
располагаются мышцы, которые действуют не только на запястный (заплюсневый)
сустав, но и на суставы пальцев.
6. Мышцы на тазовой
конечности развиты сильнее, чем на грудной, так как при передвижении животного
основная работа выполняется тазовыми конечностями.
2.9.6.
Вспомогательные
органы мышц
1.
фасции
2.
бурсы
3.
сухожилия влагалищные
4.
блоки
5.
сезамовидные кости
Фасции – это широкие пластинчатые соединительнотканные
образования, формирующие футляры для мускулатуры в целом, группы мышц, и
отдельных мышц
Функции фасции:
1.
защитная
2.
сдерживает боковое давление со стороны других мускулов на другие органы
3.
служат местом закрепления мышц
4.
создают благоприятные условия для работы мышц
Классификация фасций:
— Фасции бывают поверхностные, глубокие и специальные
(покрывают индивидуальные мышцы). Поверхностные фасции состоят из 2-х листков:
наружного и внутреннего. Между ними – подкожные мышцы (шейные,
лопаточно-плечевой мускул, и подкожные грудобрюшные мышцы). Эти мышцы не имеют
точек крепления к костной ткани, но они обеспечивают подвижность кожи. Глубокие
фасции располагаются под поверхностной. Внутренний листок глубокой фасции
выстилает изнутри стенки больших полостей организма
— Внутренний листок глубокой фасции. в больших
полостях тела имеет специальное название: внутригрудная фасция – выстилает
изнутри грудную полость, поперечная брюшная фасция: выстилают стенку брюшной
полости. наружный листок глубокой фасции в области брюшной стенки имеет в своём
составе большое количество эластических волокон, за счёт которого приобретает
жёлтую окраску и получает название жёлтая брюшная фасция она формирует
подвешивающую связку вымени.
— Специальные фасции. Они покрывают мускул в
отдельности и в отдельных участках получают специальные названия: фасция
предплечья, икроножная фасция бедра (рис. 132)
Бурсы – это мешочки, стенки которых состоят из
соединительной ткани, а полость заполняется жидкостью.
Бурсы делятся:
1. По
характеру жидкости: слизистые (подкожные, или далеко от сустава); синовиальные
2. по расположению:
подкожные, подсухожильные, подсвязочные, подмышечные.
3. по
происхождению: врождённые (постоянные); приобретённые (рис. 133).
1. Ярёмный
желоб. Является футляром наружной
ярёмной вены. Он образован мышцами плечеголовной и грудинно-челюстной.
2. Белая
мышца живота – биологический шов,
который образовался в результате апоневроза (сращения) сухожилий брюшных стенок
(наружная косая брюшная, внутренняя косая брюшная, поперечная брюшная).
3. Брюшной
пресс. Образован восьмью мышцами
брюшных стенок.
2.9.7.
Мускулатура
шеи, туловища и хвоста
Скелетная мускулатура тела
животного по выполнении функции и местоположению подразделяется на следующие
группы:
1. мышцы
головы
2. мышцы
позвоночного столба: дорсальные и вентральные
3. мышцы
грудной и брюшной стенок
4. мышцы
прикрепляющие переднюю конечность к туловищу
5. мышцы
грудной и тазовой конечности
2.9.7.1.
Дорсальные
и вентральные мышцы позвоночного столба
Мускулатура
позвоночного столба подразделяется на дорсальную и вентральную. Первая группа
мышц при двухстороннем действии разгибает позвоночный столб, а при
одностороннем – сгибает позвоночник в свою сторону. Вентральная мускулатура
сгибает позвоночный столб, подтягивает вперед таз.
К
дорсальным мышцам позвоночного столба относятся длиннейшая,
подвздошно-реберная, пластыревидная, полуостистая, многораздельная, остистая,
межпоперечные и межостистые мышцы.
Длиннейшая
мышца – m. longissimus – разделяется на длиннейшую
мышцу спины и поясницы, длиннейшую мышцу шеи и длиннейшую головы и аланта.
Длиннейшая
мышца спины и поясницы – m. longissimus dorsi et lumborum –
начинается от ости первого крестцового и остистых отростков поясничных
позвонков, маклока и подвздошного гребня, направляется краниально, заполняя
пространство между остистыми и поперечными отростками позвонков. Впереди она
истончается и заканчивается на поперечно-реберном отростке седьмого шейного
позвонка у жвачных и лошади, пятого – у свиньи, имея подсухожильную сумку.
Медиальные зубцы мышцы заканчиваются на сосцевидных (у собаки на добавочных)
отростках поясничных и последних грудных позвонков.
Длиннейшая
мышца шеи – m.
longissimus cervicis –
берет начало от поперечных отростков первых шести грудных позвонков и поперечноостистой
фасции, направляется к поперечно-реберным отросткам с 7 по 3 шейных позвонков.
Вначале мускул прикрыт длиннейшей мышцей спины и поясницы.
Длиннейшая
мышца головы и атланта – m.. longissimus capitis et atlantis –
начинается от сосцевидных отростков первых двух грудных и поперечно-реберных
отростков 7, 6, 5 позвонков и направляется к поперечному отростку эпистрофея,
крылу атланта и сосцевидному отростку каменистой кости.
Подвздошно-реберный
мускул – m. iliocostalis – начинается от крыла
подвздошной кости таза, проходит латерально от предыдущей мышцы, располагаясь
на поперечных отростках поясничных позвонков. Далее отдельными зубцами
направляется вперед, закрепляясь на латеральной поверхности ребер и поперечных
отростках с 7 по 3 шейные позвонки.
Поясничная
часть мышцы у собаки и крупного рогатого скота является самостоятельной, а у
свиньи и лошади сливается с длиннейшей мышцей поясницы.
Пластыревидная
мышца – m. splenius – начинается от канатиковой
части выйной связки, впереди крепится к крылу атланта вместе с длиннейшей
мышцей головы, затылочной кости и сосцевидному отростку.
Функция
– при одностороннем действии поворачивает шею и голову, при двустороннем -
поднимает голову.
Полуостистая
мышца головы и шеи – m. semispinalis capitis et cervicis –
начинается сухожилиями с 6 (5) по первый грудной позвонки и поперечно-реберных
отростков с 7 по 2 шейных позвонков, а заканчивается широким сухожилием на
боковой части затылочной кости и яремном отростке. Функция – поворачивает и
поднимает голову и шею.
Остистая
мышца спины и шеи – m.
spinalis dorsi et cervicis –
формируется, как самостоятельный мускул от остистых отростков 12 (10) грудных
позвонков, пучками закрепляется на остистых отростках шейных позвонков (до
4-5).
Многораздельная мышца – m. multifidus – расположена
непосредственно на позвонках в виде отдельных пучков, начиная с первого
крестцового и до 2 (3) шейных. Пучки берут начало от сосцевидных отростков и
достигают остистых отростков впереди лежащих позвонков.
Межпоперечные
мышцы – mm. intertransversarii – расположены между
поперечно-реберными отростками поясничных и шейных позвонков и участвуют в
боковых изгибах позвоночного столба.
Межостистые
мышцы – mm. interspinales – заполняют короткими пучками
пространства между остистыми отростками позвонков, помогая разгибать
позвоночный столб (рис. 134).
У
собаки и свиньи располагаются, кроме шеи, на грудных и поясничных позвонках.
К
коротким мышцам головы относятся: прямая дорсальная короткая – m. rectus capitis brevis, которая расположена
между дорсальными бугорками атланта и наружным затылочным предбугорьем; прямая
дорсальная длинная мышца – m
rectus capitis longus dorsalis, покрывающая первую и закрепляющаяся на
передней части гребня эпистрофея; косая краниальная мышца головы – m. obliquus capitis cranialis –
от переднего края крыла атланта до яремного отростка; косая каудальная мышца
головы – m. obliquus capitis caudalis –
идет от гребня эпистрофея до каудального края крыла атланта. Функции: разгибают
затылочно-атлантовый сустав и участвуют в поворотах головы.
Вентральные
мышцы позвоночного столба
К
вентральным мышцам позвоночного столба относятся длинная мышца шеи, длинная
мышца головы и прямая вентральная мышца головы, большая, малая поясничные и
квадратная мышца поясницы.
Длинная
мышца шеи – m.
longus colli –
делится на грудную и шейные части. Первая берет начало от тел первых 5 (6)
грудных позвонков, пучки волокон мышцы направлены косо вперед к поперечным
отросткам соседних позвонков. Вторая часть начинается от 7 до первого шейных
позвонков (вентральный бугорок атланта) и направлена кранио-медиально.
Длинная
мышца головы – m.
longus capitis –
лежит латеральнее предыдущей от 6 (7) поперечно-реберных отростков до мышечных
бугров тела затылочной кости.
Прямая
вентральная мышца головы – m. rectus capitis – находится на вентральной
поверхности затылочно-атлантного сустава от вентрального бугорка атланта до
мышечных бугров тела затылочной кости.
К
вентральным мышцам шеи также относятся грудинно-челюстная – m. sternomandibularis; грудинно-подъязычная – m. sternohyoideus; грудинно-сосцевидная – m. sternomastoideus и грудинно-щитовидная – m. sternothyreoideus.
Все они
начинаются на рукоятке грудины, а заканчиваются на своих объектах: 1) в области
угла нижней челюсти; 2) на члениках подъязычной кости; 3) на сосцевидном
отростке каменистой кости; 4) на щитовидном хряще гортани. Функция: помогают
опускать шею, голову, нижнюю челюсть, оттягивать гортань.
Мышцы
хвоста
Хвостовая
мышца – m. coccygeus – начинается от седалищной
ости таза, крестцово-седалищной связки и направляется к поперечным отросткам
первых 4-5 хвостовых позвонков.
Дорсальные
крестцово-хвостовые (медиальная и латеральная мышцы) – mm. sacrococcygei – расположены на
вентральной поверхности тел позвонков и поперечных отростков.
Межпоперечные
мышцы хвоста развиты слабо, в виде коротких пучков.
2.9.7.2.
Мышцы
грудных и брюшных стенок
Мышцы
грудных стенок крепятся на ребрах и грудине и способствуют изменению объема
грудной полости во время акта вдоха и выдоха. Функционально они подразделяются
на инспираторы (вдыхатели) и экспираторы (выдыхатели).
К
вдыхателям относятся: зубчатый дорсальный вдыхатель, межреберные наружные,
лестничный, прямой грудной, подниматели ребер и диафрагма. У всех инспираторов
волокна направлены каудо-вентрально.
Зубчатый
дорсальный вдыхатель – m. serratus dorsalis inspiratorius -
берет начало от пояснично-спинной фасции широким сухожилием и зубцами оканчивается на позвоночных концах с
5 по 8 ребро. Состоит из 8 зубцов у собаки, 3-6 зубцов у жвачных, 4-5 у свиньи
и 7-8 зубцов у лошади.
Лестничная
мышца – m. scalenus – состоит из надреберной (m. scalenus supracostalis), средней и
вентральной частей. Все три начинаются от поперечных отростков 5, 6, 7 шейных
позвонков. Первая оканчивается на латеральной поверхности 4-5 ребер. Вторая и
третья части оканчиваются на переднем крае первого ребра. У лошади отсутствует
надреберная часть.
Наружные
межреберные мышцы – mm. intercostales
externi
– заполняют межреберья, начинаясь в мышечных желобах ребер. Волокна направлены
дорсо-краниально на латеральную поверхность впередилежащего ребра. В области
реберных хрящей мышцы отсутствуют.
Прямая
грудная мышца – m.
rectus thoracis –
берет начало от нижней четверти первого ребра плоским сухожилием и оканчивается
на 4-5 реберном хряще, имея каудо-вентральное направление.
Подниматели
ребер – mm. levatores costarum – короткие пучки их волокон
расположены от сосцевидных отростков грудных позвонков до углов ребер, прикрыты
длиннейшей мышцей спины.
Диафрагма
–
diaphragma, m. phrenicus – самый мощный инспиратор,
разделяющий грудную и брюшную полости. Состоит из ножек, реберной части и
сухожильного центра. Правая (более мощная) и левая ножки диафрагмы крепятся под
поясничными позвонками и образуют поясничную часть. Между ножками диафрагмы
имеется отверстие для аорты и пищевода. Несколько ниже этого отверстия
расположено отверстие для каудальной полой вены.
Реберная
часть диафрагмы состоит из мышечных пучков, которые закрепляются на вентральной
поверхности ребер и грудине. Грудинная часть диафрагмы.
Сухожильный
центр – centrum tendineum –
куполообразно изгибается в сторону грудной полости, достигая уровня 6 грудного
позвонка. Спереди и сзади диафрагма покрыта серозной оболочкой грудной и
брюшной полости.
К
мышцам-выдыхателям относятся: дорсальный
зубчатый выдыхатель, внутренние межреберные, оттягиватель ребер и поперечный
грудной мускул.
Дорсальный
зубчатый выдыхатель – m. serratus dorsalis expiratorius –
берет начало от пояснично-спинной фасции, направляясь кранио-вентрально,
оканчивается на 13-10 ребрах зубцами.
Плоское мышечное брюшко формирует 3-4 зубца у крупного ротаго скота и собаки, у
овцы и свиньи – 5-6, 7 – у лошади.
Внутренние
межреберные мышцы – mm. intercostales
interni –
заполняют межреберья, прикрыты наружными межреберными мышцами. Волокна мускулов
длиной 4-
Оттягиваетль
последнего ребра – m.
retractor costae
–расположен между поперечным отростком первого поясничного позвонка и последним
ребром в виде небольшого треугольника. Направление волокон кранио-вентральное.
Поперечный
грудной мускул – m.
transversus thoracis –
лежит поперек сегментов грудины от дорсальной связки до реберных хрящей,
отдельными пучками покрывая внутреннюю поверхность последних.
Мышцы
брюшного пресса
Мышцы
живота образуют стенки брюшной полости. В их состав входят: наружный косой,
внутренний косой, поперечный и прямой грудной мускулы.
Наружный
косой мускул – m.
obliquus abdominis externus – расположен
между грудиной, реберной стенкой и тазовыми костями. Мышечная часть начинается
отдельными зубцами от грудной кости и ребер. Направление мышечных пучков
каудо-вентральное. Широкий апоневроз с таковым с другой стороны формирует белую
линию живота – linea alba,
брюшная пластинка в области таза делится на подвздошную (тазовую) и бедренную
пластинки. Подвздошная пластинка крепится от лонного бугра до маклока, имея в
своем толстый коллагеновый пучек (паховая связка).
Бедренная
пластинка оканчивается на медиальной поверхности бедра.
На
месте расхождения брюшной и тазовой пластинок у самцов маралов имеется узкая
щель – наружное отверстие пахового канала – anulus inguinalis externus для
прохождения семяпровода из придатка семенника в брюшную полость. У собак
паховое кольцо выражено у самок и
самцов.
Внутренняя
косая мышца – m.
obliquus abdominis internus –
прикрыта предыдущей, начинается от маклока и пояснично-спинной фасции,
направляется кранио-вентрально и переходит в апоневроз, который участвует в
формировании белой линии. В каудальной части апоневроза образуется щель –
внутреннее отверстие пахового канала – anulus inguinalis internus.
Поперечная
мышца живота – m.
transversus abdominis –
начинается от поперечно-реберных отростков поясничных позвонков и направляется
вертикально до мечевидного отростка реберной дуги и белой линии живота.
Принимает участие в формировании влагалища прямого брюшного мускула.
Прямая брюшная мышца – m. rectus abdominis – плоской лентой тянется от 5-6 реберных хрящей вдоль живота до лонного края, имеет 4-5 сухожильных перемычек и лежит в футляре, образованном снаружи апоневрозом наружных и внутренних косых мышц, а медиально – апоневрозами внутренних и поперечных мускулов (рис. 135).
Мышцы
живота опосредованно участвуют в акте выдоха.
2.9.8.
Мышцы
головы
Мышцы
головы у маралов подразделяются на мимические и жевательные. Первые
обеспечивают мимику, движения губ, носа, оформляют стенки ротовой полости. Вторые
обеспечивают движение нижней челюсти при перетирании пищи и захвате корма
Мимическая
мускулатура
К
мимическим мышцам относят круговой мускул рта – m. orbicularis oris, расположенный в основе
губ. Мускул имеет наружную и внутреннюю (ротовую) поверхности.
Подниматель
верхней губы – m.
levator labii dorsalis
начинается в области лицевого бугра, направляется вперед к верхней губе, где и
заканчивается. Функция – поднимает верхнюю губу и расширяет ноздри.
Клыковой
мускул – m. caninus – берет начало там же, где
и предыдущий мускул и заканчивается на крыле носа. Функция – расширяет ноздри.
Опускатель
верхней губы – m.
depressor labii dorsalis –
начинается в области лицевого бугра и заканчивается в области угла рта и
латеральной части верхней губы, сливаясь с предыдущими мышцами. Функция –
опускает верхнюю губу.
Носогубной
подниматель – m.
levator nasolabialis –
представлен тонкой косой пластинкой, начинающейся от лобной фасции основания носовых
костей, и направляется к верхней губе и верхушке носа. Оканчивается в углу
ноздри и верхней губе. Функция – поднимает верхнюю губу и расширяет ноздрю.
Скуловая
мышца – m. zygomaticus – лентовидной формы,
расположена подкожно на латеральной части морды. Начинается на фасции большого
жевательного мускула, оканчивается в области угла рта. Функция – оттягивает
угол рта («улыбка»).
Опускатель
нижней губы – m.
depressor labii ventralis –
начинается от фасции массетера, идет по щечной поверхности нижней челюсти и
заканчивается в нижней губе. Функция – помогает опускать нижнюю губу.
Щечная
мышца – m. buccalis – расположена в основе
щеки, снаружи прикрыта кожей, внутри – слизистой оболочкой ротовой полости.
Состоит из двух пластинок – наружной, имеющей вертикально-косое направление, и
внутренней, имеющей горизонтальный ход волокон. Обе пластинки начинаются в
области угла рта, вплетаясь в круговой мускул. Функция – образует стенки
ротовой полости, удерживая пищу.
Верхняя
и нижняя резцовые мышцы – m. incisivus dorsalis et ventralis –
начинаются соответственно в области резцовой кости и резцового края нижней
челюсти, а заканчиваются в верхней и нижней губе. Функция – помогают поднимать
верхнюю губу и оттягивать угол рта книзу.
Подбородочная
мышца – m. mentalis – начинается пучками от
резцовой части нижней челюсти и заканчивается в коже подбородка. Функция –
оттягивает нижнюю губу, сморщивая подбородок.
Поперечный
мускул носа – m.
transversus nasi –
начинается на резцовой кости, содержит поверхностные и глубокие пучки и
оканчивается на медиальном крыле носа. Функция – расширяет ноздри.
Боковой
мускул носа – m.
lateralis nasi –
развит относительно хорошо и состоит у маралов из медиального и латерального
расширителя. Функция – расширяет ноздри.
Жевательная
мускулатура
К
жевательной группе относятся большой жевательный мускул, височный, крыловой и
двубрюшный мускулы. Эти мышцы участвуют в движении нижней челюсти.
Большая
жевательная мышца – m.
masseter –
расположена на боковой поверхности ветви нижней челюсти, заполняя одноименную
ямку. Имеет два слоя: наружный – начинается в области лицевого бугра и
скулового гребня, направляется каудо-вентрально, закрепляясь позади сосудистой
вырезки нижней челюсти. Внутренний слой имеет вертикальное направление волокон.
Двубрюшная
мышца – m. digastricus – начинается от яремного
отростка формирует ростральное и каудальное брюшко, которое опускается по
каудальному краю ветви нижней челюсти. Ростральное брюшко заканчивается на
ветви нижней челюсти. Функция – опускает челюсть.
Височная
мышца – m. temporalis – расположена в височной
ямке черепа и заканчивается на венечном отростке нижней челюсти. Функция –
поднимает нижнюю челюсть, смыкая рот.
Крыловидная
мышца – m. pterygoideus – состоит из двух слоев.
Верхний край ее крепится в области крыловидного отростка клиновидной кости.
Медиальный пласт короткий и веерообразно направлен в сторону суставного
отростка нижней челюсти. Латеральный пласт заполняет крыловидную ямку нижней
челюсти, закрепляясь на ее каудальном крае. Функция – выдвигает нижнюю челюсть
вперед и смещает в сторону (рис. 136).
Мышцы
языка и подъязычной кости
Затылочно-подъязычный
мускул – m. hyooccipitalis – берет начало от яремного отростка
и оканчивается на мышечном отростке стилогиоида. На яремном отростке можно
выделить мышечную и сухожильную части.
Шилоподъязычный
мускул – m. stylohyoideus – разделен на две части,
которые начинаются на мышечном отростке стилогиоида. Длина латерального
сухожилия 15-
Рожковоподъязычный
мускул – m. keratohyoideus – лежит виде тонкой
пластинки между латеральной поверхностью тиреогиоида, кератогиоидом и
эпигиоидом.
Поперечный
межчелюстной мускул – m. transversus mandibulae – состоит из
язычно-челюстного и подъязычно-челюстного. Первый начинается от подбородочного угла
нижней челюсти и до первого премоляра. Подъязычно-челюстной мускул развит лучше
предыдущего и простирается от уровня второго премоляра каудально до базигиоида
и тиреогиоида, имея два направления мышечных волокон.
Шилоязычный
мускул – m. styloglossus – начинается мышечной
частью от латеральной поверхности стилогиоида и оканчивается на боковой
поверхности языка вплоть до его верхушки.
Хрящеязычный
мускул – m. chondroglossus – хорошо выражен, прикрыт
шилоязычным мускулом. Берет начало на дистальном конце стилогиоида, занимая всю
его каудальную поверхность, далее направляется вперед на боковую поверхность
языка.
Рожковоязычный
мускул – m. keratoglossus – берет начало от
кератогиоида, эпигиоида и частично стилогиоида, направляется к корню языка,
вплетаясь в подбородочноязычный мускул.
Подбородочноязычный
мускул – m. genioglossus – самый мoщный из описанных, начинается от
подбородочного угла нижней челюсти коротким сухожилием, рядом с одноименным
мускулом другой стороны и распределяется в верхушке, теле и корне языка,
отдавая пучок волокон к надгортаннику.
Язычноподъязычный
мускул – m. hyoglossus – начинается длинной
полосой от латеральной поверхности базигиоида и нижней части тиреогиоида,
оканчиваясь в теле языка.
Подбородочноподъязычный
мускул – m. geniohyoideus – начинается сухожилием
длинной 40-
Подъязычнощитовидный
мускул – m. hyothyreoideus – берет начало от задней
части пластинки щитовидного хряща и заканчивается на базигиоиде и тиреогиоиде.
У марала он закрепляется только на дистальном отделе базигиоида поверхностной и
глубокой частями.
2.9.9.
Мышцы,
прикрепляющие грудную конечность к туловищу
Трапецевидная
мышца – m. trapezius – имеет шейную и грудную
части, берет начало от надостистой связки со 2 шейного до 11-12 грудного
сегментов. Заканчивается на ости лопатки, причем шейная часть имеет
каудо-вентральное направление волокон и частично сливается с плечеголовным
мускулом. Грудная часть имеет кранио-вентральное направление волокон. Функция:
вращает лопатку.
Ромбовидная
мышца – m. rhomboideus – расположена под предыдущей
мышцей. Шейная часть узкая, берет начало от второго шейного до второго грудного
позвонков, грудная часть со 2 по 5 грудные позвонки. Обе части оканчиваются на
медиальной поверхности лопаточного хряща. Функция, как у трапециевидного
мускула.
Зубчатая
вентральная мышца – m.
serratus ventralis –
начинаеися веерообразно от зубчатой поверхности лопатки и делится на шейную и
грудную части. Шейная часть заканчивается на поперечно-реберных отростках со 2
по 7 шейных позвонков, грудная – с 1 по 5 ребра. Функция: подвешивает туловище
между передними конечностями.
Плечеголовная
мышца – m. brachiocephalicus – расположена от плечевого
сустава и до затылка. Начинается от латерального гребня плечевой кости, ниже
дельтовидной шероховатости рядом с дельтовидной мышцей и делится на
ключично-затылочную и ключично-сосцевидную части. Ключично-затылочная (поверхностная)
часть берет начало от ключичной полоски (рудимент ключицы) и оканчивается на
затылочном гребне и канатиковой части выйной связки. Ключично-сосцевидная часть
мускула (глубокая) идет от ключичных полосок и заканчивается на мышечном бугре
тела затылочной кости вместе с длинным мускулом головы.
Грудинно-челюстная
мышца – m. sternomandibularis – в виде ленты расположена
от рукоятки грудины и до угла нижней челюсти, прикрыта m. masseter.
Грудинно-сосцевидная
мышца – m. sternomastoideus – начинается в области
рукоятки грудины, идет под предыдущем мускулом и у атланта сливается с
сухожилиями ключично-сосцевидного и длинного мускула головы.
Атланто-акромиальная
мышцы (плече-атлантная) – m. atlantoacromialis – лежит между нижним концом
лопатки и крылом атланта, прикрыта в большей части ключично-затылочной мышцей.
Функция – подтягивает нижний угол лопатки вперед.
Широчайшая
мышца – m. latissimus dorsi – берет начало от
пояснично-спинной фасции, направляется кранио-вентрально к круглой
шероховатости плечевой кости, прикрывая боковую грудную стенку. Функция: при
движении животного выносит конечность назад, а при стоячем положении
подтягивает тело вперед.
Поверхностная
грудная мышца – m.
pectoralis superficialis –
состоит из плечевой и предплечной частей. Первая начинается от рукоятки грудины
и первых 4-5 ее члеников, направляется к гребню большого бугра плечевой кости,
где и заканчивается на фасции медиальной поверхности предплечья. Предплечная
часть берет начало от 1-6 реберных хрящей и заканчивается на фасции медиальной
поверхности предплечья.
Глубокая
грудная мышца – m.
pectoralis profundus –
имеет плечевую и предлопаточную части. Плечевая часть берет начало от брюшной
фасции, тела грудной кости и реберных хрящей (с 4 по 8), направляется к малому
бугру плечевой кости, где закрепляется. Плоское сухожилие мышцы может достигать
большого бугра плечевой кости, где соединяется с сухожилиями двуглавого и
предостного мускулов. Лопаточная часть, более слабая, берет начало от первых
4-х реберных хрящей и оканчивается на переднем крае предостного мускула (рис. 137)
2.9.10.
Мышцы
передней конечности
Статический аппарат грудной
конечности:
Разновидность сил действующих
на грудную конечность проходит через основание лопатки, через середину
локтевого и запястного суставов, и опускается позади третьей фаланги пальцев.
При действии этой силы плечевой сустав стремится к флексии. Статику плечевого
сустава обеспечивает сухожилие двуглавого мускула плеча. Статику плечевого
сустава и запястного сустава обеспечивает лучевой разгибатель запястного
сустава и фиброзный тяж.
Статику запястного сустава и
суставов пальцев обеспечивают 3 тяжа:
1. Глубокий
тяж. Представлен 3-им межкостным мускулом (закрепляется дистальным концом на
сезамовидных костях, а связки сезамовидных костей закреплены на костях первой
фаланги). Обеспечивает статику первой фаланги.
2. Средний
тяж. Представлен сухожилием глубокого пальцевого сгибателя
3. Наружный
тяж. Представлен поверхностным сгибателем пальцев (рис. 138).
2.9.10.1.
Мышцы
плечевого и локтевого суставов
Предостная
мышца – m. supraspinatus – заполняет предостную ямку
лопатки и заканчивается плоскими сухожилиями на латеральных и медиальных буграх
плечевой кости, пропуская между ножками сухожилие двуглавого мускула. Функция:
экстензор плечевого сустава.
Заостная
мышца – m. infraspinatus – заполняет одноименную
ямку лопатки, идет вдоль ее ости и оканчивается под латеральным бугром плечевой
кости, имея подсухожильную бурсу. Функция: абдуктор и фиксатор.
Дельтовидная
мышца – m. deltоideus – берет начало от фасции лопатки,
закрывает с поверхности заостную мышцу и оканчивается между акромионом и
дельтовидной шероховатостью гребня плечевой кости (акромиальная часть), а также
на дельтовидной шероховатости (лопаточная часть). Функция: фиксатор и абдуктор
плечевого сустава.
Подлопаточная
мышца – m. subscapularis – заполняет ямку от ее
зубчатой шероховатости и заканчивается на медиальном мышечном бугре плечевой
кости. Функция: аддуктор.
Большая
круглая мышца – m.
teres major –
начинается от каудального угла лопатки и длинной лентой тянется до круглой
шероховатости плечевой кости, замыкая треугольник «лопатка – плечевая кость –
круглый мускул». Функция: флексор и аддуктор.
Малая
круглая мышца – m.
teres minor –
расположена от шейки лопатки до гребня большого бугра плечевой кости. Прикрыта
заостной и дельтовидной мышцами. Функция: флексор и аддуктор.
Коракоидно-плечевая
мышца – m. coracobrachialis – начинается от коракоидного
отростка лопатки плоским сухожилием, имеющим подсухожильную бурсу, проходит
между предостным и подлопаточным мускулом и оканчивается на медиальной стороне
плечевой кости от медиального бугра до ее середины. Функция: экстензор
плечевого сустава.
Мышцы,
действующие на локтевой сустав
Двуглавая
мышца плеча – m.
biceps brachii –
начинается от бугра лопатки и коракоидного отростка мощным сухожилием, которое
лежит в межбугорковом желобе плечевой кости, где имеет под собой синовиальную
бурсу. Далее она переходит в веретенообразное мышечное брюшко. Заканчивается
мускул на лучевой шероховатости и медиальной поверхности локтевой кости. От
нижней трети мускула отделяется сухожильная ножка – lacertus fibrosus, идущая к лучевому
разгибателю запястья. Функция: флексор локтевого сустава, помогает лучевому
разгибателю.
Внутренняя
плечевая мышца – m.
brachialis internus –
берет начало от задней поверхности шейки плечевой кости между латеральной и медиальной
головками трицепса, переходит на латеральную поверхность плеча и заканчивается
на лучевой кости ниже места крепления двуглавого мускула. Функция: флексор
локтевого сустава.
Трехглавая
мышца – triceps
brachii
– расположена между лопаткой, плечевой костью и локтевым бугром. Мускул имеет
три головки: длинную, латеральную и медиальную.
Длинная
головка – caput longum –
начинается от каудального края лопатки и заканчивается на локтевом бугре, имея
под собой бурсу.
Латеральная
головка – caput laterale
–
берет начало от шейки и латерального бугра плечевой кости и заканчивается на
локтевом бугре.
Медиальная
головка – caput mediale –
начинается двумя частями от медиального гребня и круглой шероховатости плечевой
кости. На уровне середины плечевой кости обе части срастаются и заканчиваются
на локтевом бугре вместе с длинной головкой. Функция: экстензор локтевого
сустава.
Локтевая
мышца – m. anconaeus – слабо развита, начинается
от мыщелков плечевой кости вокруг локтевой ямки и оканчивается на латеральной
поверхности локтевого отростка.
Напрягатель
фасции предплечья – m.
tensor fascii antebrachii –
узкой полоской идет вдоль медиальной поверхности длинной головки трехглавого
мускула и оканчивается на фасции предплечья и локтевом бугре. Его начало – от
каудального угла лопатки (рис. 139).
Функция:
экстензор.
2.9.10.2.
Мышцы
запястного сустава и пальцев
Лучевой
разгибатель запястья – m. extensor carpi radialis – расположен на
дорсо-латеральной поверхности предплечья. Берет начало от латерального
надмыщелка плечевой кости и заканчивается на специальной шероховатости
дорсальной поверхности пясти. Сухожилие мускула заключено в синовиальное
влагалище.
Локтевой разгибатель запястья – m. extensor carpi ulnaris – начинается от
разгибателя надмыщелка плечевой кости и заканчивается на добавочной кости
запястья. Он оформляет задний контур предплечья. Функция – флексор.
Длинный
абдуктор большого пальца – m. abductor policis longus – слабо развит, лежит
на дорсальной поверхности нижней трети лучевой кости, а виде плоского
треугольной формы мускула. Его сухожилие перекрещивает лучевой разгибатель и
закрепляется на медиальной поверхности головки пястной кости.
Лучевой
сгибатель запястья – m. flexor carpi radialis – расположен на
медиальной поверхности предплечья. Начинается от медиального надмыщелка
плечевой кости, заканчивается на пальмарной поверхности проксимальной части
пясти. Между лучевым сгибателем и лучевой костью формируется срединный желоб,
где расположены сосуды и нервы.
Локтевой
сгибатель запястья – m. flexor carpi ulnaris – расположен на
медиально-пальмарной поверхности предплечья. Начинается двумя головками –
плечевой и локтевой. Первая берет начало от медиального надмыщелка плечевой
кости, вторая – от локтевого бугра. Обе головки, сливаясь, оканчиваются на
добавочной кости запястья.
Общий
пальцевый разгибатель – m. extensor digitorum communis -
расположен на латеральной поверхности предплечья и состоит из двух головок.
Одна, более мощная, начинается от латерального надмыщелка плечевой кости и
фасции лучевого разгибателя запястья. Другая, более слабая, головка берет
начало от латеральной связки локтевого сустава. Мускул окружен синовиальным
влагалищем.
Поверхностный
пальцевый сгибатель – m. flexor digitorum superficialis – состоит
из двух мышечных частей, которые берут начало от медиального надмыщелка
плечевой кости, лежит между локтевым сгибателем запястья и глубоким сгибателем
пальцев с пальмарной поверхности предплечья.
Оба
сухожилия находятся в синовиальном влагалище. В области запястья они
фиксированы поперечной пальмарной связкой. На пясти сухожилия становятся
плоскими, прикрывают сверху таковые глубокого сгибателя пальцев и оканчивается
на вторых фалангах 3 и 4 пальцев.
Глубокий
сгибатель пальцев – m.
flexor digitorum profundus –
состоит из плечевой, локтевой и лучевой головок, которые берут начало от
медиального надмыщелка плечевой кости, локтевого отростка и головки луча.
Мышца
расположена в желобе между костями предплечья, проходит через запястье, образуя
желобок для сухожилия поверхностного сгибателя пальцев. В области пясти
сухожилие глубокого сгибателя проходит между сухожильными ножками
поверхностного сгибателя и оканчивается на сгибательной поверхности копытцевых
костей 3 и 4 пальцев.
Межкостная
мышца – m. interossеus medius – в виде широкого плоского
сухожилия лежит на пальмарной поверхности пясти и служит подвешивающей связкой
для сезамовидных костей.
Короткие
сгибатели пальцев, как мышцы, не выражены, а представляют
собой сухожильные пучки к длинным сгибателям (рис. 140).
2.9.11.
Мышцы
тазовой конечности
Статический аппарат тазовой
конечности.
При действии силы на тазовую
конечность коленный сустав и заплюсневый сустав стремятся согнуться, а суставы
пальцев – разогнутся. Статику коленного сустава обеспечивают 2 тяжа:
1.
Представлен 3-им малоберцовым мускулом
2.
Представлен икроножной мышцей и поверхностным сгибателем пальцев
— Статику сустава пальцев тазовой конечности
обеспечивает сухожилия мышц как и на грудной конечности (рис. 141)
2.9.11.1.
Мышцы
тазобедренного и коленного суставов
Мускулатура
тазовой конечности включает мышцы, действующие на тазобедренный, коленный,
заплюсневый и пальцевые суставы.
К
мышцам тазобедренного сустава относятся: ягодичные, двуглавая, грушевидная,
квадратная, двойничная, полусухожильная, полуперепончатая,
подвздошно-поясничная, напрягатель широкой фасции бедра, портняжная,
гребешковая, приводящая, запирательные.
Поверхностная
ягодичная мышца у марала, как самостоятельная, не выражена.
Каудальная часть ее слилась с седалищной головкой двуглавого мускула бедра,
образовав его крестцовую головку.
Средняя
ягодичная мышца – m.
gluteus medius –
заполняет поверхность крыла подвздошной
кости таза и оканчивается на большом вертеле бедренной кости, прикрывая
глубокую ягодичную мышцу. Функция: экстензор тазобедренного сустава.
Глубокая
ягодичная мышца – m.
gluteus profundus –
лежит на ягодичной поверхности крыла подвздошной кости под средним ягодичным
мускулом. Оканчивается, как и предыдущая мышца, на большом вертеле. Функция:
экстензор тазобедренного сустава.
Грушевидная
мышца – m. piriformis – лежит впереди от средней
ягодичной мышцы и закрепляется на большом вертеле ниже предыдущей. Функция:
экстензор тазобедренного сустава.
Подвздошно-поясничная
мышца – m. iliopsoas – входит в состав конечного
участка большой поясничной мышцы и двух частей поясничной мышцы. Первая описана
ранее, как вентральная мышца позвоночного столба. Подвздошная мышца начинается
на теле подвздошной кости и все три части заканчиваются на малом вертеле
бедренной кости. Функция: флексор тазобедренного сустава, подтягивает таз
вперед.
Напрягатель
широкой фасции бедра – m. tensor fasciae latae – оформляет передний
контур бедра, лежит непосредственно под кожей. Начинается от нижней части
маклока, заканчивается плоским сухожилием на медиальной поверхности коленного
сустава. Функция: флексор тазобедренного и экстензор коленного суставов.
Квадратная
мышца – m. quadratus femoris – расположена со стороны
плантарной поверхности бедра от седалищной ости и до верхней трети бедренной
кости. Функция: экстензор тазобедренного сустава.
Двойничная
мышца – m. gemellus – начинается от малой
седалищной вырезки таза и заканчивается в вертлужной ямке бедренной кости между
сухожилиями запирательных мышц. Функция: супинатор.
Наружная
и внутренняя запирательные мышцы – mm. оbturatorii externi et interni –
начинаются в области запертого отверстия таза – внутренняя – с дорсальной,
наружная – от вентральной поверхности. Обе заканчиваются в вертлужной ямке
бедренной кости. Функция: супинаторы.
Портняжная
мышца – m. sartorius – лежит в виде ленты,
начинаясь от фасции подвздошно-поясничной мышцы и поясничного бугорка тела
подвздошной кости таза. Заканчивается мышца на медиальной стороне фасции
коленного сустава. Функция: флексор тазобедренного и экстензор коленного
суставов.
Гребешковая
мышца – m. pectineus – в виде веретенообразного
брюшка расположена на медиальной стороне бедра от подвздошно-лонного возвышения
таза до малого вертела бедренной кости. Функция: аддуктор и флексор
тазобедренного сустава.
Стройная
мышца – m. gracilis – лежит на медиальной
стороне бедра в виде плоской, довольно толстой пластины, сзади срастаясь
сухожильной ножкой с полусухожильной и полуперепончатой мышцами. Начинается от
тазового шва и заканчивается на медиальной стороне фасции голени вдоль гребня
большеберцовой кости. Функция: аддуктор конечности.
Приводящая
мышца – m. adductor femoris – хорошо развита, довольно
толстая, прикрыта стройным мускулом, начинается от тазового сращения и
заканчивается на середине диафиза бедренной кости. Функция: аддуктор
конечности.
Полусухожильная
мышца – m. semitendinosus – лежит в заднебедренной
области, впереди частично прикрыта двуглавым мускулом. Начинается от
седалищного бугра и крестцовой фасции, опускается по каудально-медиальной части
бедра и закрепляется на пяточном бугре, фасции голени и медиальной поверхности
гребня большеберцовой кости. Функция: экстензор тазобедренного сустава, флексор
коленного и экстензор заплюсневого суставов.
Полуперепончатая
мышца – m. semimembranosus – в виде ленты лежит
медиальнее предыдущей, начинаясь от нижнего отрога седалищного бугра, сливаясь
здесь с полусухожильной мышцей. Далее достигает по медиальной стороне бедра
коленного сустава, где оканчивается широким сухожилием на фасции.
Двуглавая
мышца бедра – m.
biceps femoris –
срастаясь у марала с поверхностной ягодичной мышцей, образует
ягодично-двуглавую мышцу (m. gluteobiceps), как у северного оленя. Мускул
имеет вид широкой пластинки, начинается крестцовой головкой от
пояснично-крестцовой фасции и седалищной головкой от соответствующего бугра,
оканчивается тремя головками: коленной – на латеральной части коленной чашки, голенной – на гребне
большеберцовой кости и пяточной – на пяточном бугре. Функция: экстензор
тазобедренного, заплюсневого и флексор коленного суставов, супинирует
конечность.
Мышцы,
действующие на коленный сустав
Четырехглавая
мышца бедра – m.
quadriceps femoris – самый
мощный мускул тазовой конечности. Имеет четыре головки: латеральную,
медиальную, прямую и добавочную.
Прямая
головка – m. rectus femoris – самая массивная среди
других. Начинается от тела подвздошной кости над суставной впадиной таза и
направляется вдоль дорсальной поверхности бедренной кости до коленной чашечки,
прикрывая добавочную головку и срастаясь с ней в нижней трети бедра.
Добавочная
головка – vastus accessorius
берет начало от проксимальной части тела бедренной кости.
Латеральная
головка – vactus lateralis –
начинается от нижней части большого вертела, медиальная – от медиальной
поверхности шейки бедренной кости. Все головки оканчиваются на коленной чашке.
Функция: экстензор коленного сустава и флексор тазобедренного (прямая головка).
Подколенная
мышца – m. popliteus – начинается от специальной
ямки латерального мыщелка бедренной кости и заканчивается в области плантарной
поверхности шейки большеберцовой кости. Функция: флексор коленного сустава (рис. 142).
2.9.11.2.
Мышцы
заплюсневого сустава и пальцев
К
мышцам, действующим на заплюсневый сустав, относятся: передняя большеберцовая,
третья малоберцовая, задняя большеберцовая, трехглавая мышца голени, длинная
малоберцовая.
Трехглавая
мышца голени – m.
triceps surae –
состоит из латеральной, медиальной головок и пяточной мышцы. Латеральная и
медиальная головки образуют икроножный мускул. Начинаются в
области соответствующих надмыщелков бедренной кости, на середине голени
сливаются в мощное пяточное (Ахиллово) сухожилие, которое заканчивается на
пяточном бугре, имея под собой сухожильную бурсу.
Пяточная
мышца – m. soleus – берет начало от
малоберцового отростка, идет косо к латеральной головке икроножного мускула,
где с ней и сливается на уровне середины голени. Функция: экстензор заплюсны и
флексор коленного сустава.
Передняя
большеберцовая мышца – m. tibialis cranialis – находится непосредственно
на большеберцовой кости, прикрыта третьим малоберцовым мускулом. Начинается от
проксимальной части большеберцовой кости и заканчивается на дорсальной
поверхности проксимального конца плюсневой кости. Функция: флексор заплюсневого
сустава.
Третья
малоберцовая мышца – m. peroneus tertus – образует переднюю
поверхность голени, начинается от разгибательной ямки латерального надмыщелка
бедренной кости и заканчивается на проксимальной части плюсны. Функция: флексор
заплюсны.
Длинная
малоберцовая мышца – m. peroneus longus – лежит рядом с предыдущей,
начинаясь от малоберцового отростка, тонким сухожилием заканчивается на
латеро-дорсальной поверхности плюсневой кости. Функция: флексор заплюсны.
Мышцы
пальцев тазовой конечности
К
ним относятся: длинный разгибатель пальцев, специальный разгибатель III и IV пальцев, короткий разгибатель пальцев,
поверхностные и глубокие сгибатели пальцев.
Длинный
разгибатель пальцев – m. extensor digitorum longus –
берет начало вместе с малоберцовой третьей мышцей, проходит через заплюсневый
сустав, удерживаясь сухожильной перемычкой, следует по дорсальной поверхности
плюсны, оканчиваясь сухожилиями на третьих фалангах пальцев.
Специальный
разгибатель III и IV пальцев – m. extensor digitorum III et IV –
берет начало от латерального надмыщелка бедренной кости, лежит рядом с длинным
разгибателем и заканчивается на дистальных фалангах III и IV пальцев.
Короткие
разгибатели, как мышцы, у жвачных не выражены. Их заменяют сухожильные пучки к
окружающим мышцам.
Поверхностный
сгибатель пальцев - m.
flexor digitorum pedis sublimis –
расположен между латеральной и медиальной
головками икроножного мускула, берет начало от задней поверхности
дистального эпифиза бедренной кости, лежит на плантарной поверхности голени. Он
одевает сверху пяточный бугор, опускается на плюсну и оканчивается на
дистальных фалангах всех пальцев.
Глубокий
сгибатель пальцев – m. flexor digitorum profundus –
состоит из поверхностной, глубокой и медиальной головок. Первые две лежат
латерально и начинаются от проксимальной части большеберцовой кости и
малоберцового отростка, а медиальная головка – от подколенной вырезки между
мыщелками большеберцовой кости. На плантарной поверхности большеберцовой кости
все головки переходят в мощное сухожилие, которое перекидывается через
держатель таранной кости, далее идет по плантарной поверхности плюсны, под
сухожилием поверхностного сгибателя. Оканчивается мускул на дистальных фалангах
пальцев.
Межкостная
мышца, как на передней конечности, представлена мощным сухожильным тяжем (рис. 143)
3.
Кожа
и ее производные
3.1.
Общая
морфофункциональная характеристика кожного покрова и его производных
Кожный
покров – integumentum
commune –
выполняет многообразные функции и, в первую очередь, интегрирует все органы,
представляя громадное рецепторное поле, реагирующее на все, даже самые
незначительные воздействия внешней среды, в которой обитает животное. Кожа
защищает организм от механических, физических, химических и биологических
воздействий, участвует в обмене веществ и терморегуляции организма. Последнее
особенно важно для животных, обитающих в условиях Крайнего Севера и
высокогорья. Высокая плотность пуховых волос, а также наличие воздуха в остевых
волосах значительно увеличивает теплоизоляционные свойства волосяного покрова.
У северного оленя, например, по данным Сегаль А.Н. (1983), Овсова А.С. (1993),
Кушнир А.В., Соколова А.Я., Гречкиной Л.И. (2003) тканевая теплоизоляция при
понижении температуры внешней среды возрастает за счет снижения температуры
кожи на выступающих участках тела (конечности. уши). Так, при температуре
окружающей среды 0ºС температура кожи голени составляет +30ºС, а при
температуре окружающей среды -40ºС – всего +9º +11ºС, в то же
время температура кожи на боковых участках тела была постоянной (+36ºС).
Важное
значение при этом имеет пиломоторная реакция (состояние волосяного покрова). По
данным Lentz C., Hart J. (1960), эта реакция приводит к
уменьшению теплоотдачи на 35%.
Вопросам
морфологии кожного покрова у различных видов животных посвящено довольно
большое количество работ. Так, Непесова А.Г. (1970), Приселкова Д.О. (1973),
Прасолова Л.А. (1980), Петрищев Б.И. и Лебенгарц Л.З. (1986) исследовали
морфологию кожи и кожных желез у крупного рогатого скота. Макеев В.В. (1972),
Панфилова Я.Т. и Тинякова Р.С. (1986) – у коз оренбургской пуховой породы.
Гистоморфологию
кожи северного оленя описали Боль Б.К. и Николаевский А.Д. (1932), Браун А.А. и
Островская П.И. (1933), Акаевский А.И. (1939), Ефимов А.Е. (1939, 1940),
Шумилов М.Р. и Ильин П.А. (1977).
В
своих работах И.С. Ржаницына и Н.Д. Овчаренко (1984-1986) дают описание
различных участков кожного покрова и его производных (панты и хвостовая железа)
у маралов и пятнистых оленей. Липовских А.А. (1984) исследовал нервные элементы
губ маралов различных возрастных групп.
3.2.
Функции
кожи
— Защитная;
— Терморегулирующая;
— Выделительная;
— Секреторная;
— Рецепторная;
— Эндокринная;
— Депонирующая;
— Коммуникационная.
3.3.
Строение
кожи и источники ее происхождения
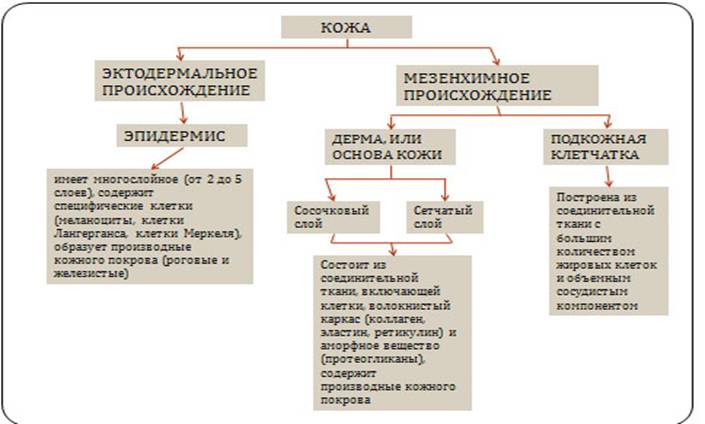
ЭПИДЕРМИС
Это поверхностный относительно тонкий слой участков кожи,
покрытых волосами, и более толстый в безволосых участках. Эпидермис –
многослойный плоский ороговевающий эпителий (рис. 145).
ДЕРМА
Это соединительнотканная часть кожи, состоящая из
аморфного вещества, содержащего протеогликаны, коллагеновые волокна и их пучки,
эластические волокна с их участием сети, а также различные виды клеток:
фибробласты-фиброциты, макрофаги, тучные клетки, липоциты, плазмоциты и др.
Сосочковый
слой расположен непосредственно под эпидермисом и представляет
собой совокупность соединительнотканных сосочков, находящихся между гребнями
эпидермиса и имеющих разную высоту и форму. Этот слой наиболее выражен на
безволосых участках кожи животных или в коже с редким волосяным покровом
(щетина у свиней). В коже с густым волосяным покровом (шерсть у овец)
механические и другие воздействия на организм опосредованы через волосы. В
такой коже эпидермис тонкий и его граница с дермой более ровная, т. е. сосочки
выражены слабо. Так как в этом слое находятся корни волос, то у овец его
нередко называют пилярным от латин. Pili – волосы.
Сосочковый слой состоит из рыхлой соединительной ткани, в
которой преобладают клетки и аморфный компонент. Аморфное вещество здесь имеет
большое значение в матаболизме, транспорте питательных веществ от
гемокапилляров к клеткам сосочкового слоя и к клеткам эпидермиса, а также в
перемещении продуктов метаболизма в обратном направлении. По этой причине,
исходя из функционального назначения, сосчковый
слой нередко называют трофическим.
В участках кожи с повышенной чувствительностью (кожа губ,
хоботок у свиней, молочная железа и др.) в соединительной ткани сосочков
находятся многочисленные чувствительные нервные окончания. Среди них есть
свободные – это терморецепторы, и болевые рецйепторы, а также инкапсулированные
тельца Мейснера – механорецепторы, реагирующие на смещение кожи при
прикосновении к ней , т.е. осязание.
Сеть эластических волокон сосочкового слоя в комплексе с
эластическими сетями других зон дермы создает общий эластический каркас кожи.
СЕТЧАТЫЙ
СЛОЙ
Располагается в дерме
глубже сосочкового и состоит из плотной неоформленной соединительной ткани. В
этом слое коллагеновые волокна и образованные из них пучки ориентированы в
разных направлениях, вследствие чего в дерме формируются своеобразные
трехмерные сети, которые в каждом участке кожи наилучшим образом отвечают на
направленные на них механические воздействия и обьсепечивают прочность данного
участка.
В кожном покрове дорсальных областей туловища (спина и
круп крупных животных) кожа мало растяжима, и конструкция переплетения пучков
коллагеновых волокон в этом слое имеет упорядоченный ромбический вид. На
вентральной стороне туловища кожа часто подвергается растяжению (при
беременности, при увеличении объема живота после принятия пищи), и в этой
области пучки коллагеновых волокон более тонкие и они располагаются
преимущественно горизонтально.
В микроструктуре сетчатого слоя дермы обнаружены и
существенные породные различия. Например, у овец грубошерстной романовской породы выявлено усложненное переплетение
коллагеновых волокон. У овец тонкорунных
пород пучки коллагеновых волокон более тонкие и преобладает их
горизонтальная ориентация. Вокруг пучков коллагеновых волокон сформированы сети
эластических волокон, направление которых соответствует ориентации таких
пучков.
ГИПОДЕРМА.
По функциональному назначению – это связующий слой с
соединительной тканью глубже расположенных органов произвольного
движения. Построена из соединительной ткани с большим количеством
жировых клеток и объемным сосудистым компонентом (рис.
146)
3.4.
Кровоснабжение
кожи
— Собственно
кожные и мышечно-кожные артерии обеспечивают кровоснабжение кожи.
— Имеется
поверхностная и глубокая сосудистые сети. Первая расположена на границе
сосочковой и ретикулярной дермы. Из кожного сплетения между подкожной
клетчаткой и дермой отдельные артериолы идут вертикально к поверхности и
разветвляются у подножия сосочкового слоя, образуя субпапиллярное сплетение,
которое обеспечивает питание эпидермиса;
— Вторая
сосудистая сеть располагается на границе дермы и гиподермы. Она соединяется с
поверхностной сетью вертикально идущими к ней сосудами. В гиподерму отходят
сосуды, питающие жировые дольки;
— Наличие
большого числа артерио-венозных анастомозов, играющих важную роль в
терморегуляции (рис. 147).
3.5.
Внутрикожные
железы
К
внутрикожным железам у млекопитающих относят потовые и сальные железы; молочные
и модифицированные (пахучие – мускусные железы у кабарги, бобра, кролика),
межкопытные железы овец, оленей, железы анального региона (всеядные и хищные). (рис.
148)
Особенности потовых
желез у домашних животных
|
Вид
животного |
Степень
развития и расположения на теле |
|
Лошадь |
Развиты
сильно, расположены на вентральной поверхности груди и живота, вентральной и
боковой поверхности шеи, в области промежности паха |
|
Корова |
Хорошо
развиты на морде и шее (в остальных местах развиты слабо) |
|
Свинья |
Разбросаны
по всей поверхности тела, хорошо развиты на морде и шее |
|
Овца |
Развиты
сильно, особенно на дорсальной поверхности туловища и тазовых конечностях,
кожи мошонки, области ануса и препуция |
|
Коза |
Очень
слабо развиты |
|
Собака |
Развиты
слабо, в большом количестве расположены в области спины |
|
Кошка |
Очень
слабо развиты, сосредоточены только на безволосой поверхности опорных
участков лап |
|
Кролик
|
Очень
слабо развиты, имеются только в области губ |
Функции
потовых желез
} Защиты кожи от
высыхания;
} Создание на
поверхности кислой среды;
} Поддержание
гомеостаза воды и солей;
} Экскреция некоторых
веществ: хлоридов, мочевины;
} Терморегуляция;
} У овец потовый секрет
смешивается с кожным салом и образует защитную смазку - жиропот
3.5.1.
Мерокриновые
потовые железы
представляют
собой трубочки диаметром 30-100 мкм, один конец (внутрикожный) которых замкнут,
а другой свободно открывается на поверхности эпидермиса, не имея связи с
корнями волос. В железе по ее длине, строению стенки и функции выделяют
концевой секреторный отдел и выводной проток. Секреторный отдел у овец и у крс
имеет форму извилистой трубочки, у свиней и лошадей форму клубка. Такие
концевые отделы располагаются, как правило, в глубокой части дермы, на границе
с гиподермой. В коже грубошерстный овец они находятся на уровне волосяных
луковиц остевых волос, в коже свиней на уровне луковиц щетины, в коже
тонкорунных овец – несколько глубже уровня волосяных луковиц (рис. 149)
3.5.2.
Сальные
потовые железы
в
эмбриогенезе – как часть волосяного зачатка – связаны с корнями волос. В коже сосков молочной железы, носогубного
зеркала – самостоятельно (рис. 150).
К
производным кожного покрова относятся также молочные железы.
Вымя – uber – расположено у коров расположено в
срамной области гипогастрия. Вымя кроме сосков покрыто волосом. В коже сосков
отсутствуют волосы, сальные и потовые железы, эпидермис утолщен.
Под
кожей вымени располагается поверхностная фасция в виже беловатой прочной пластинки.
Под ней лежит собственная оболочка долей вымени.
Вымя
разделено на левую и правую половины. Между ними по сагиттальной плоскости
расположена подвешивающая связка вымени из эластичной ткани.
В
вымени различают основание, тело и соски. В соске различают основание, тело и
кончик.
Формы
вымени:
Чашеобразное
– тело вымени округлое, высота значительная, соски расставлены широко, доли
незначительно различаются по размерам;
Ваннообразное
– доли хорошо развиты, тело значительной высоты, вытянутое, краниальный край
приближен к пупочной области мезогастрия;
Плоское
– отличается от чашеобразного меньшей высотой;
Козье
– характеризуется конусообразным тело, сближенными по расположению сосками,
высота задних долезначительно превышает высоту передних;
Примитивное
– небольшого размера с небольшими сближенными сосками.
Система,
выводящее молоко состоит из:
Молочных
протоков – ductus lactiferi;
Молочного
синуса – sinus lactiferi;
Крупные
молочные протоки вливаются в цистерну (молочный синус).
Выводящая
система заканчивается сосковым протоком, в нем имеется внутреннее сосковое и
наружное отверстие. Сфинктеры (вокруг сосокового протока) регулируют
прохождение молока.
У
маралух расположено между бедрами в задней части брюшной стенки. По средней
сагиттальной линии оно разделено бороздой на правую и левую половины, каждая из
которых имеет передний и задний молочные холмы. Передние холмы шириной до
Каждая
из долей молочной железы имеет самостоятельную систему выводных протоков,
открывающихся в полость соответствующей молочной цистерны. Молочные ходы и
выводные протоки, идущие от секреторных отделов железы, открываются в стенке
добавочных полостей, реже в стенке центральной полости железистой цистерны.
Подвешивающая
связка делит молочную железу на правую и левую половины и состоит из двух
сросшихся соединительнотканных листков, в которых много эластических волокон.
Поверхностная
фасция вымени построена из циркулярно направленных коллагеновых волокон, среди
которых встречаются островки жировой ткани. В прослойках фасции проходят
крупные кровеносные сосуды и нервные пучки поверхностного нервного сплетения.
Поверхностная фасция переходит без границ в глубокую фасцию, от которой в толщу
железы отходят прослойки, делящие паренхиму на доли и дольки.
В
соединительнотканных прослойках залегают сосуды и нервные пучки, идущие к
секреторным отделам. Междольковые выводные протоки выстланы двуслойным
цилиндрическим эпителием.
Паренхима
вымени представлена альвеолами и выводными протоками, объединенными в
секреторные дольки. Границы долек слабо обозначены тонкими
соединительно-тканными прослойками.
3.5.3.
Апокриновые
потовые железы
развиваются и располагаются
в определенных участках кожного покрова. У овец, антилоп и газелей они
находятся вблизи глаз, у грузынов – в паховой, прианальной и препуциальной
областях, у оленей и лис – в корне хвоста. Орудие защиты, другим – узнавание и
нахождение себе подобных, а также полового привлечения особей друг к другу.
Хвостовая
железа хорошо развита у маралов и располагается у основания
хвоста под эпидермисом, имеет зернистый вид, размеры ее 3-
3.6.
Носовое
зеркало
Особый
интерес представляют дерматоглифические рисунки носогубного зеркала у
маралов. По данным Ю.В. Аржанковой (2001, 2002), которая описала дерматоглифику
носогубного зеркала у крупного рогатого скота в связи с происхождением и
породной продуктивностью, можно выделить несколько видов рисунков (рис. 152).
Для носогубного зеркала маралов характерна сильная
пигментация, поэтому поверхность темного цвета. Кожные
валики в дорсальной и центральной частях носогубного зеркала 5-6-угольной
формы, а в вентральной – эллипсоидной. Кожные борозды располагаются в различных
направлениях, одиночно, с асимметричным рисунком левой и правой сторон.
Выявлено два типа дерматоглифа: «малина» и «каменная брусчатка»:
1) «малина» – валики сильно выпуклые, расположены кучно, по 5-9 штук, межваликовые кожные борозды очень глубокие, между «ягодами» по всей поверхности носогубного зеркала вкраплены волосяные островки, их до 35 штук. Такой тип дерматоглифа присущ примерно 65 % животных;
2)
«каменная брусчатка» – валики умерено выпуклые, расположены равномерно, в основном одиночно, межваликовые борозды
глубокие волосяных островков до 19 штук.
Носогубное зеркало маралов васкуляризируется
двумя сосудистыми магистралями: артерией верхней губы и дорсальной ветвью
подглазничной артерии. Причём эти артерии ассиметричные: с левой стороны имеют
несколько больший диаметр, чем с правой. Артерия верхней губы, имеющая диаметр
в среднем 3 мм, отходит от лицевой артерии, делает волнообразный изгиб в
области 1-2 премоляра, направляется дорсо-медиально и подходит к вентральной
части носогубного зеркала. Здесь от нее отходят в виде кустиков более мелкие
сосуды диаметром в среднем 1 мм, концы которых истончаются и сплетаются в
плотную сосудистую сеть. Эта сеть своими апекальными концами между ноздрями
переходит из вентральной части в дорсальную вертикальными тяжами и опять
образует сосудистую сеть. Дорсальная часть носогубного зеркала
васкуляризируется дорсальной ветвью подглазничной артерии, имеющей средний
диаметр 1 мм, которая ответвляется от основной подглазничной артерии, имеющей
диаметр в среднем 2 мм, на выходе из
подглазничного отверстия и волнообразно направляется сначала рострально, а
затем дорсо-назально и подходит к носогубному зеркалу. В данной области от
дорсальной ветви подглазничной артерии отходят мелкие сосудистые веточки и
вплетаются в сосудистую сеть носогубного зеркала (рис.
153).
3.7.
Строение
волоса
В каждом волосе по его длине
различают часть, находящуюся над поверхностью кожи – стержень и часть, расположенную
на различной глубине внутри кожи – корень волоса. Внутрикожный конец волоса
утолщен и получил название луковицы волоса. Со стороны основания луковица имеет
утолщение – волосяной сосочек, в рыхлой соединительной ткани которого имеются
кровеносные капилляры и рецепторные нервные окончания. Все эпителиальные части
корня волоса окружены соединительно тканной оболочкой – волосяной сумкой (рис. 154).
Стержень – мозговое вещество,
корковое вещество и кутикула.
Мозговое вещество находится в
центральной осевой части стержня. В нем находится небольшое количество грнаул
пигмента и много воздушгых полостей. Развит в остевых и отсутствует в пуховых.
У северного оленя 98% толщины стержня. Воздух - плохой проводник тепла, но волос
легко ломается.
Корковое вещество – в остевых волосах и
щетине. По длине образуются роговые чешуйки – содержится наибольшее количество
пигмента.
Кутикула – чешуйчатый слой :
три типа кутикул: кольцевидный, некольцевидный и мостовидный.
Категории волоса:
— Направляющие
(диам.около 100 мкм) – поддерживают более тонкие пуховые волосы, придают
красивый внешний вид;
— Остевые – несколько
короче и тоньше направляющих, луковицы располагаются глубже пуховых;
— Пуховые образуют
более глубокий ярус волосяного покрова. Они более тонкие и короткие, луковицы
располагаются в дерме поверхностно;
— Промежуточные
характерны для полутонкой шерсти;
— Щетина не имеет
сердцевины, но сильно развит кортекс;
— Вибриссы
(осязательные волосы)
3.8.
Копыто,
копытце, коготь
Копытце – unguicula, как производное кожи,
покрывает третьи фаланги основных и боковых пальцев. Копытце состоит из
копытцевой каймы, венчика, стенки и копытцевой подошвы (рис.
155).
Копытцевая
кайма представляет узкую безволосую полоску шириной 2-
Основа
кожи каймы построена из тонких и длинных сосочков и богата кровеносными
сосудами. Подкожный слой состоит из соединительной ткани с наличием
коллагеновых волокон.
Венчик
копытца выступает в виде небольшого валика. Эпидермис венчика расположен на
длинных сосочках дермы и продуцирует трубчатый рог копытца, опускающийся по
мере роста до подошвы. В венчике имеется подкожный слой.
Стенка
копытца состоит из эпидермиса и собственно кожи без подкожного слоя. Основа
кожи копыцевой стенки образует многочисленные листочки высотой до
Копыто – ungula. Расположено на пальце лошади.
В стенке роговой капсулы различают краниальную переднюю
зацепную часть и по бокам от нее медиальную и латеральную части боковой стенки,
которые затем переходят пальмарно или плантарно в заворотные части стенки – pars inflexa lateralis et medialis,
обращенные к подошве копыта. Углы поворота боковой стенки к подошве азывают
латеральными и медиальными плантарными или пальмарными краями или заворотными углами.
Венчик основы кожи занимает меньшую, чем в копытцах,
часть от высоты стенки.
В стенке роговой капсулы копыта находятся более толстые
слои трубчатого и рога. Белая зона
хорошо обозначена, по которой вбивают гвозди, закрепляющие подкову (рис. 156)
Механика копыта может быть нарушена в результате:
• недостатка движения;
• плохого и неправильного питания;
• плохого грунта;
• плохих условий содержания;
• холода и жары;
• излишней влажности;
• повреждения роговых трубочек;
• потери глазури;
• ну и конечно, в результате ковки, которая нарушает естественную функцию
расширения копыта.
Коготь – unguicula – имеет форму крючка с
заостренной вершиной. В нем различают три части:
Кайма на
границе с кожей в виде валика с желобом, покрывающий костный валик и желоб
когтевой кости;
Спинковая
и боковые части стенки когтя;
Подошва
когтя.
Поверхность когтя покрыта роговой капсулой, под которой
лежат производящий слой эпидермиса и основа кожи – дермис. Подкожный слой
соединительной ткани есть только под основой кожи каймы в области валика и
проксимальной части когтевой кости. Подкожный слой отсутствует в области
средины и вершины когтевой кости. Здесь дермис срастается с надкостницей
фаланги.
Когти под влиянием мышц могут втягиваться в желоб валика
и выдвигаться из него.
3.9.
Рога
Рога
–
cornu –
расположены на роговых отростках лобных костей, покрыты роговой капсулой, образованной эпидермисом рога. Под эпидермисом находится основа кожи рога с
красноватыми сосочками благодаря значительному количеству в них кровеносных
сосудов. Сосочки создают значительную поверхность для питания производящего слоя эпидермиса,
образующего роговую капсулу рога.
В
роге различают верхушку – apex cornus, основание рога – basis cornus и восковицу – epiceras (кайма). В основании рога восковица переходит
в кожу.
Верхушка
рога имеет только роговой слой, поэтому ее можно спиливать безболезненно.
Панты (от
китайского Фун-ту) – это неокостеневшие рога маралов, которые вырастают только
у самцов и используются как ценное сырье в Тибетской медицине. Длина пантов
может достигать
Для
марала характерным является следующее расположение отостков: надглазничный –
отходит сразу от «коронки»; «ледяной» – следует за предыдущим; средний – на
некотором расстоянии от «ледяного». Далее появляются пятый и шестой отростки,
расположенные в одной плоскости. Панты марала подвержены высокой степени
индивидуальной изменчивости.
Первые
рога (шпильки) появляются у самцов в 12-13 месяцев. Спад шпилек происходит во
второй половине апреля – первой половине мая.